
Abstrak
Padang rumput merupakan dasar pakan ternak ruminansia; namun, daya cerna hijauan menurun pada musim kemarau. Dalam penelitian ini, tujuannya adalah untuk mengidentifikasi isolat jamur terpilih dari saluran pencernaan domba Santa Inês yang sehat dan untuk mengevaluasi daya cerna in vitro jerami Urochloa decumbens (UD) yang diinokulasi dengan jamur ini. Analisis sekuens daerah ITS rDNA dan fragmen gen pengkode β-tubulin memungkinkan identifikasi isolat O45M1 sebagai Aspergillus terreus . Isolat B13M2 diidentifikasi sebagai Trichoderma longibrachiatum dengan mempertimbangkan analisis sekuens daerah ITS dan gen pengkode faktor elongasi 1-α. Koefisien kecernaan bahan kering (DMD) in vitro dan kecernaan serat deterjen netral (NDFD) dari UD lignifikasi dievaluasi dalam cairan rumen yang dikumpulkan dalam tiga periode 21 hari. Cairan rumen diinokulasi dengan kaldu Sabouraud steril (kontrol), dengan A. terreus (O45 M1), dengan T. longibrachiatum (B13 M2), atau dengan campuran jamur ini. DMD jerami lebih tinggi ( p < .01, uji Fisher LSD) ketika diinokulasi dengan isolat B13M2 (47,31%) dibandingkan dengan kontrol (35,69%) untuk tiga periode pengumpulan cairan rumen. Kehadiran jamur miselium terdeteksi dalam sampel media rumen selama tiga kali inkubasi (0, 24 dan 72 jam) dalam simulator rumen. Selain itu, setelah pencernaan asam, pada 72 jam inkubasi, populasi jamur miselium secara signifikan lebih tinggi ketika cairan rumen diinokulasi dengan isolat B13M2 ( p < .05, uji Fisher LSD), yang menunjukkan ketahanan. Penambahan B13M2 meningkatkan DMD dari UD lignifikasi, menunjukkan potensi untuk pengembangan aditif probiotik atau mikroba untuk ruminansia.
1. PENDAHULUAN
Di wilayah tropis, kualitas hijauan menurun drastis selama musim kemarau, yang disebabkan oleh proses fisiologis dan lignifikasi dinding sel (Wang et al., 2016 , Sousa et al., 2023 ). Genus Urochloa telah dibudidayakan secara luas di daerah-daerah ini karena menunjukkan toleransi kekeringan dan tumbuh baik di tanah dengan kesuburan sedang hingga rendah, serta menyediakan produksi massal yang tinggi di wilayah tropis dan subtropis (Ferreira et al., 2021 ; Pessoa-Filho et al., 2017 ). Namun, ketika hijauan ini menunjukkan kualitas nutrisi yang rendah, mengoptimalkan kecernaannya melalui penggunaan aditif mikroba, probiotik atau prebiotik sangat penting untuk meningkatkan produktivitas ruminansia yang dibesarkan dalam kondisi seperti itu. Suplementasi jamur eksogen dan/atau ekstrak dan enzimnya pada diet ruminansia dapat meningkatkan penggunaan serat berkualitas rendah dan meningkatkan asupan bahan kering, serta produksi ruminansia (López-Aguirre et al., 2016 ; Pittaluga et al., 2025 ). Namun, terdapat ketidakkonsistenan dalam hasil penggunaan aditif ini, terutama pada jenis galur jamur, yang sering kali tidak bertahan hidup dengan baik di lingkungan rumen (López-Aguirre et al., 2016 ; Uwineza et al., 2024 ). Bahasa Indonesia: Dalam penelitian kami sebelumnya, keberadaan genus Aspergillus , Paecilomyces , Acremonium , Trichoderma , Malbranchea , dan Onychocola terdeteksi dalam sampel dari saluran pencernaan (DT) domba yang dibesarkan di padang rumput tropis (Freitas et al., 2012 ; Martins Júnior et al., 2023 ). Selain itu, isolat Aspergillus dan Trichoderma yang dipilih dari rumen sapi telah menunjukkan degradasi struktur vegetal kompleks oleh produksi enzim hidrolitik yang ekspresif (Abrão et al., 2017 ; Abrão et al., 2021 ; Duarte et al., 2021 ). Dalam penelitian sebelumnya, satu isolat Aspergillus sp. dan satu isolat Trichoderma sp. dari DT domba menunjukkan aktivitas karboksimetilselulase sebesar 350,39 dan 139,2 μmol mL −1 , masing-masing, pada 72 jam fermentasi U. decumbens yang mengalami lignifikasi (Freitas, 2018 ). Kami berhipotesis bahwa penyertaan jamur terpilih dari saluran pencernaan domba (DT), atau campuran jamur ini, dapat meningkatkan degradasi makanan berserat tinggi, sehingga menghasilkan peningkatan keuntungan ekonomi bagi ternak yang dibesarkan di feedlot atau di padang rumput. Dalam penelitian ini, tujuannya adalah untuk melakukan identifikasi molekuler dengan analisis filogenetik dari dua jamur terpilih dari DT domba dan untuk mengevaluasi kecernaan in vitro jerami U. decumbens yang diinokulasi dengan jamur ini.
2 BAHAN DAN METODE
2.1 Isolasi jamur
Isolat jamur yang dievaluasi dikumpulkan dari percobaan sebelumnya yang disetujui oleh Komite Etika Eksperimen Hewan dari Universidade Federal de Minas Gerais (protokol n° 128/2013), yang diatur oleh Dewan Pengawasan Eksperimen Hewan Nasional Brasil. Jamur Trichoderma sp. (B13 M2) dan Aspergillus sp. (O45 M1) diisolasi dari ampul rektal domba Santa Inês berusia 6 hingga 7 bulan yang diberi makan, masing-masing, dengan Panicum maximum cv. Tanzania (Freitas et al., 2012 ) atau Cynodon dactylon cv. Vaqueiro di utara Minas Gerais, Brasil, selama musim kemarau. Isolat ini adalah yang paling sering, menunjukkan populasi >106 unit pembentuk koloni (CFU) per gram feses dan menunjukkan produksi enzim degradasi dinding sel tanaman yang unggul. Selain itu, jamur ini diidentifikasi menurut metode yang dijelaskan oleh De Hoog et al. ( 2000 ), dengan kultur mikro dan analisis karakteristik morfologi mikro. Dalam penelitian sebelumnya, isolat ini dikulturkan dalam medium C, yang mengandung selulosa mikrokristalin sebagai satu-satunya sumber karbon, untuk evaluasi aktivitas selulolitik (CA) dan menunjukkan indeks CA >2, yang menunjukkan pertumbuhan yang baik saat menggunakan polisakarida ini sebagai sumber karbon tunggal (Martins Júnior et al., 2023 ).
2.1.1 Identifikasi molekuler
Jamur tersebut ditumbuhkan dalam agar Sabouraud (Acumedia, Lansing, Michigan, AS) selama 7 hari, dan DNA diekstraksi menggunakan glukanase (Glucanex; Novo Nordisk Ferment Ltd., Dittingen, Swiss) dalam satu ekstraksi fenol diikuti oleh dua ekstraksi fenol/kloroform menurut Neuhauser et al. ( 2009 ). Wilayah internal transcribed spacer (ITS) rDNA diperkuat dari DNA yang diekstraksi melalui reaksi berantai polimerase (PCR) menggunakan primer ITS1 (TCCGTAGGTGAACCTGCGG) dan ITS4 (TCCTCCGCTTATTGATATGC), menurut Toth et al. ( 2013 ). Selain itu, fragmen gen yang mengkode protein β-tubulin (β-tub) diperkuat dengan penggunaan Bt2a (GGTAACCAAATCGGTGCTGCTTTC) dan Bt2b (ACCCTCAGTGTAGTGACCCTTGGC) (Glass & Donaldson, 1995 ) dan fragmen gen faktor elongasi (EF-1α) diperkuat menggunakan primer EF-728 M (CATYGAGAAGTTCGAGAAGG) dan EF2 (GGARGTACCAGTSATCATGTT) (Samson et al., 2014 ). Reaksi PCR, dengan volume akhir 50 μL, berisi 5 μL buffer 10 ×, 5 μL MgCl 25 mM, 1 μL dNTP 10 mM, 2 μL setiap primer pada 10 μM, 2,5 μL DMSO pada 20%, 2 μL DNA pada 150 ng, 0,5 μL DNA polimerase Taq (Sinapse, São Paulo, BR) dan 30 μL H 2 O ultramurni.
Amplifikasi dilakukan dengan denaturasi awal pada suhu 94°C selama 5 menit, diikuti oleh 35 siklus denaturasi pada suhu 94°C selama 1 menit, annealing primer ITS pada suhu 58°C selama 1 menit 20 detik, primer β-tub pada suhu 55°C selama 1 menit, dan primer EF-1αa pada suhu 57°C selama 1 menit, ekstensi pada suhu 72°C selama 1 menit dan 30 detik, diikuti oleh ekstensi akhir pada suhu 72°C selama 5 menit. Untuk pemurnian produk PCR, 11,25 μL EDTA (125 mmol/L) dan 135 μL etanol absolut ditambahkan ke tabung yang berisi campuran reaksi. Campuran diinkubasi selama 15 menit pada suhu 28°C dan kemudian disentrifugasi pada 20.215 g selama 25 menit untuk mengendapkan amplikon. Supernatan dibuang dan 120 μL etanol 70% (v/v) ditambahkan ke tabung mikrosentrifugasi. Tabung-tabung ini dihomogenkan dengan cara dibalik, disentrifugasi pada 20.215 g selama 10 menit, dan supernatan dibuang lagi. Setelah penguapan total etanol sisa, produk disuspensikan kembali dalam 10 μL air bebas nuklease. Produk yang diamplifikasi diukur dengan NanoDrop 1000ND (Thermo Fisher Scientific, Wilmington, Amerika Serikat), dan konsentrasinya disesuaikan menjadi 100 ng μL −1 untuk digunakan dalam reaksi sekuensing.
Reaksi sekuensing dilakukan pada pelat 96-sumur menggunakan volume akhir 10 μL. 20 ng produk amplifikasi yang dimurnikan ditambahkan ke dalam sumur bersama dengan 1,6 μL buffer reaksi, 0,8 μL BigDye® Terminator v3.1 Ready Reaction Mix (AppliedBiosystems São Paulo, Brasil), 1 μL inisiator amplifikasi yang sama pada konsentrasi 5 μmol L −1 dan air. Amplifikasi dilakukan dengan denaturasi pada suhu 96°C selama 1 menit, diikuti oleh 35 siklus denaturasi pada suhu 96°C selama 15 detik, annealing primer pada suhu 50°C selama 15 detik, dan ekstensi pada suhu 60°C selama 4 menit. Untuk presipitasi reaksi sekuensing, volume 2,5 μL EDTA (125 mmol/L) dan 30 μL etanol (95%) ditambahkan ke setiap sumur. Setelah 15 menit, sentrifugasi dilakukan pada 2500 g selama 45 menit; supernatan dibuang, dan 30 μL etanol (70% v/v) ditambahkan ke setiap sumur. Sentrifugasi lebih lanjut dilakukan pada 2500 g selama 15 menit; supernatan dibuang, dan pelat dibiarkan pada suhu ruangan hingga etanol benar-benar kering (White et al., 1990 ).
Produk pemurnian disuspensikan dalam Hi-Di™ formamide dan diurutkan dalam sequencer ABI Prism 3100 (Applied Biosystems, Foster City, California, AS). Urutan yang diperoleh dipangkas untuk kualitas menggunakan perangkat lunak Bioedit Versi 7.2.5 dan Asparargim ( http://asparagin.cenargen.embrapa.br/phph/ ) untuk mengecualikan daerah berkualitas rendah (Skor Phred <20) dan untuk perakitan contig, urutan nukleotida yang diperoleh diedit dan dibandingkan dengan urutan yang disimpan dalam basis data GenBank ( http://www.ncbi.nlm.nih.gov/ ), menggunakan program Blast N (Altschul et al., 1997 ). Untuk dianggap milik spesies tertentu, isolat harus menunjukkan 97% kesamaan dengan yang lain yang sudah disimpan di GenBank (Stackebrandt & Goebel, 1994 ).
2.1.2 Analisis filogenetik
Urutan daerah ITS dan fragmen gen pengkode β-tubulin dianalisis untuk isolat O45M1, dan urutan daerah ITS dan fragmen gen pengkode faktor elongasi 1-α dianalisis untuk isolat B13M2. Analisis individual dilakukan dengan urutan terkonkatenasi yang sesuai milik spesies dalam genus. Untuk genus Trichoderma , urutan spesies dari bagian Harzianum , Viride , Virens , dan Longibrachiatum dianalisis, dan urutan spesies Hypomyces aurantius dan Sphaerostilbella aureonitens digunakan sebagai kelompok eksternal. Untuk genus Aspergillus , spesies dari bagian Fumigati , Nigri , Flavi , Circumdati , Terrei , Clavati , Usti , Candidi , Nidulantes dan Restricti dianalisis, dan spesies Talaromyces piceae dan Talaromyces bacillisporus dipilih sebagai kelompok eksternal.
Untuk setiap analisis, sekuens diselaraskan menggunakan metode Clustal (Thompson et al., 1994 ) dengan penyesuaian manual untuk perbaikan visual bila perlu dan dirangkai menjadi satu set data menggunakan program MEGA X versi 10.1 (BETA) (Kumar et al., 2018 ) Penyelarasan dilakukan dengan menyertakan sekuens klade dan bagian utamanya yang disimpan di GenBank, dengan memprioritaskan, bila memungkinkan, sekuens dari koleksi kultur atau publikasi terindeks untuk memastikan identifikasi yang akurat. Kriteria Informasi Akaike digunakan untuk mengidentifikasi model evolusi yang paling tepat untuk setiap set data. Kedua set data yang dirangkai dibentuk menggunakan Metode Kemungkinan Maksimum (ML), yang dilakukan dengan RAxML v. 8.2.4 (Stamatakis, 2006 ). Model penggantian nukleotida GTRGAMMA digunakan selama pembuatan 1000 replikasi bootstrap (Felsenstein, 1985 ). Analisis dilakukan menggunakan platform CIPRES Science Gateway (Miller et al., 2010 ). Semua urutan nukleotida disimpan di GenBank dan diberi nomor akses MK817113.1 dan MT355642 ( isolat Aspergillus sp. O45M1) dan MK817114.1 dan MT355643 ( isolat Trichoderma sp. B13 M2).
2.2 Makanan Ternak
Jerami Urochloa decumbens (UD) yang dibungkus setelah jatuhnya benih dibeli secara komersial di wilayah Montes Claros, Negara Bagian Minas Gerais, Brasil, selama musim kemarau (Maret hingga Oktober). Tiga sub-sampel jerami ini yang menunjukkan 95,38% DM dikumpulkan untuk evaluasi bromatologi (AOAC, 2010 ), yang menunjukkan komposisi berikut berdasarkan 100% bahan kering (g 100 g −1 ): protein kasar = 3,06 ± 0,23; protein tidak larut dalam deterjen netral = 21,57 ± 3,2; protein tidak larut dalam deterjen asam = 20,59 ± 3,3; serat deterjen netral (NDF) = 82,26 ± 7,3; serat deterjen asam (ADF) = 53,04 ± 7,2; lignin = 7,50 ± 0,9; ekstrak halus = 1,02 ± 0,16; total gula = 3,02 ± 0,45; abu = 5,89 ± 0,67; kalsium = 0,14 ± 0,09; fosfor = 0,10 ± 0,003; kalium = 0,58 ± 0,07; magnesium = 0,20 ± 0,04; sulfur = 0,06 ± 0,0001; selulosa = 45,54 ± 3,37; hemiselulosa = 29,22 ± 3,02; karbohidrat tak berserat = 8,43 ± 0,59.
2.3 Pengumpulan cairan rumen
Untuk uji kecernaan in vitro, dua domba Santa Inês dewasa dengan fistula rumen, masing-masing dengan berat sekitar 65 kg, digunakan sebagai donor rumen. Domba jantan ditempatkan di teluk selama 15 hari, yang 14 hari di antaranya untuk adaptasi terhadap diet yang diformulasikan untuk memenuhi kebutuhan nutrisi domba dewasa (NRC, 2007 ), yang mengandung 60% jerami UD dan 40% konsentrat dan campuran mineral. Pada hari ke-15, cairan rumen dari dua hewan dikumpulkan, dihomogenisasi, disaring dan diangkut dalam botol termal tertutup. Pengumpulan dilakukan pada pagi hari sebelum makanan diberikan. Tiga pengumpulan cairan rumen dilakukan dengan interval 21 hari untuk memungkinkan rekonstitusi mikrobiota rumen (Dirksen, 1993 ). Analisis makroskopis dilakukan segera setelah pengumpulan dalam tabung yang berisi 5 mL sampel cairan. Warna, bau, viskositas dan potensi reduksi metilen biru (MBRP) pada konsentrasi 0,03% dievaluasi.
pH cairan rumen diperkirakan menggunakan potensiometer digital (Dirksen, 1993 ). Analisis fisikokimia cairan rumen yang dikumpulkan pada tiga waktu pengambilan sampel dianggap serupa mengingat MBRP <6 menit dan baunya aromatik untuk semua kelompok hewan yang dievaluasi dalam percobaan ini. Potensi hidrogenionik (pH) rata-rata cairan rumen tetap mendekati netral (7,03). Semua prosedur dengan hewan telah disetujui oleh Komite Etik dalam Eksperimen Hewan Universidade Federal de Minas Gerais, Brasil (no 128/2013 ).
2.4 Kecernaan in vitro serat bahan kering dan serat deterjen netral jerami Urochloa decumbens
Percobaan kecernaan in vitro dirancang sebagai faktorial 4×3, membandingkan efek dari empat jenis inokulum dalam tiga periode pengumpulan cairan rumen, dengan delapan ulangan masing-masing. Kantong Filter Bag F57 (Ankon®) (5,5 cm × 5,5 cm) yang dikeringkan pada suhu 55°C selama 24 jam digunakan. Kantong kosong ditimbang dan 0,5 g jerami UD yang distandarisasi pada 1 mm (panjang dan lebar) dimasukkan ke dalamnya. Delapan kantong Ankon F-57 digunakan di setiap ruang fermentasi, yang berisi 100 mL kaldu Sabouraud steril (kontrol), dengan Aspergillus sp. (O45 M1), dengan Trichoderma sp. (B13 M2) atau dengan campuran 50 mL setiap inokulum jamur. Delapan kantong kosong yang tidak berisi isolat jamur dan tidak ada sampel jerami juga disertakan untuk setiap ruang. Penentuan in vitro dilakukan dengan dan tanpa penambahan jamur atau campuran kedua jamur.
Percobaan dilakukan dalam tiga periode dengan selang waktu 21 hari antar pengumpulan cairan rumen.
Untuk menyiapkan inokulum, jamur dikultur dalam kaldu Sabouraud pada suhu 39°C selama 48 jam dan distandarisasi pada konsentrasi sekitar 107 CFU /mL. Koefisien kecernaan bahan kering in vitro (DMIVD) dan kecernaan serat deterjen netral (NDFD) jerami hijauan, menurut metodologi yang dijelaskan oleh Tilley dan Terry ( 1963 ) dan modifikasi yang dijelaskan oleh Holden ( 1999 ), dievaluasi menggunakan simulator rumen (TE-150, Tecnal Scientific Equipment, São Paulo, Brasil). Peralatan ini terdiri dari empat ruang kaca (2500 mL) untuk inkubasi pada suhu 39°C dengan laju rotasi lima putaran per menit (rpm).
Sampel tetap bersentuhan dengan masing-masing inokulum dan cairan rumen ditambah larutan penyangga McDougall selama 48 jam pada suhu 39,5°C. Selanjutnya, 8 g pepsin dan 40 mL HCl 6 N ditambahkan untuk mensimulasikan pencernaan kimiawi, dan sampel kemudian diinkubasi selama 24 jam lagi. Pada akhir proses ini, kantong dicuci dengan air suling, dikeringkan, dan ditimbang.
Untuk penentuan NDFD, kantong yang telah dicuci dianalisis menggunakan simulator rumen dalam larutan NDF (selama 1 jam pada suhu 105°C), diikuti dengan tiga kali pencucian dengan air suling selama 10 menit dan aseton selama 15 menit, lalu didehidrasi dalam oven udara paksa pada suhu 55°C selama 24 jam dan 2 jam dalam oven pada suhu 105°C. Penentuan DMIVD diperoleh dengan rumus berikut (Ankom, 2014 ):
![]()
di mana W1 = berat kantong kosong; W2 = berat sampel; W3 = berat kantong + residu setelah inkubasi; C1 = koreksi sampel putih (berat akhir tanur kering/berat awal); DM = bahan kering.
2.5 Kuantifikasi jamur selama analisis kecernaan in vitro
Selama uji kecernaan in vitro dalam stimulan rumen pada periode pengumpulan ketiga, lima sampel media rumen dikumpulkan pada waktu 0 jam, 24 jam dan juga 72 jam, setelah penambahan asam klorida 6 N, menggunakan spuit steril 10 mL. Pengenceran desimal serial (10 −1 , 10 −2 , 10 −3 , 10 −4 dan 10 −5 ) dilakukan, dan 100 μL alikuot diinokulasi ke dalam plat steril yang berisi media agar Sabouraud. Inokulum dihomogenisasi dengan loop Drigalski steril, dan plat diinkubasi pada suhu 39°C dan dipantau untuk pertumbuhan koloni jamur hingga 21 hari (Abrão et al., 2014 ; Hoog et al., 2000 ).
2.6 Analisis statistik
Distribusi normal dan homogenitas varians dinilai menggunakan uji Lilliefors dan Bartlett, masing-masing. Percobaan kecernaan in vitro dirancang sebagai 3 × 2 × 2, membandingkan tiga periode pengumpulan cairan rumen dan ada/tidaknya A. terreus dan ada/tidaknya T. longibrachiatum sebagai inokulum, dengan delapan kali ulangan masing-masing. Data kecernaan relatif diserahkan untuk analisis varians, dan rata-rata yang paling menarik secara biologis dibandingkan dengan uji Fisher LSD.
Kuantifikasi populasi jamur dalam media rumen dievaluasi selama uji kecernaan in vitro untuk periode pengumpulan ketiga. Transformasi log 10 ( x + 10) dilakukan (di mana x adalah unit pembentuk koloni, CFU) dan konsentrasi jamur dievaluasi dalam 3 × 2 × 2 dengan membandingkan efek tiga kali inkubasi (0, 24 dan 72 jam) dan ada/tidaknya A. terreus dan ada/tidaknya T. longibrachiatum sebagai inokulum. Rata-rata yang paling menarik secara biologis dibandingkan dengan uji Fisher LSD. Semua prosedur dilakukan dengan menggunakan System for Statistical Analysis, SAEG, Versi 9.1 dengan menggunakan tingkat signifikansi 5% ( p = .05).
3 HASIL
3.1 Karakterisasi molekular isolat jamur
Dalam penelitian ini, analisis sekuens daerah ITS rDNA dan fragmen gen pengkode β-tubulin mengidentifikasi isolat O45M1 sebagai Aspergillus terreus . Isolat B13M2 diidentifikasi sebagai Trichoderma longibrachiatum , dengan mempertimbangkan analisis sekuens daerah ITS dan gen pengkode faktor pemanjangan 1-α (Tabel 1 ).
| Identifikasi terisolasi | Bahasa Inggris: Primer | Identifikasi | [Tanpa setoran – GenBank] | Jumlah pasangan basa yang dianalisis | Kesamaan (%) | Hasil BLAST [n° menurut GenBank] |
|---|---|---|---|---|---|---|
| O45M1 | DIA | Aspergillus terreus | MK817113.1 | 583 | Rp 99.000 | A. terreus Ef6 [KX 816799.1] |
| bak mandi β | A. terreus | MT355642 | 553 | 100 | A. terreus [SQUCC13244] | |
| Pesawat B13M2 | DIA | Trichoderma longibrachiatum | MK817114.1 | 615 | Rp 99.000 | T.longibrachiatum SKF-3 [KX463453.1] |
| EF 1-α | T. longibrachiatum | MT355643 | 529 | 100 | T . longibrachiatum [LES F009] |
Saat mengevaluasi profil filogenetik untuk kedua isolat, klade yang terbentuk mendukung identifikasi taksonomi molekuler. Pohon untuk isolat B13 M2 dari genus Trichoderma (Gambar 1 ) menunjukkan jamur ini sebagai anggota genus Trichoderma dan mengonfirmasi bahwa penyelarasan sekuens isolat domba ini berada dalam klade Longibrachiatum, khususnya dengan dua sekuens T. longibrachiatum lainnya .
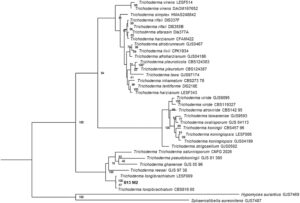
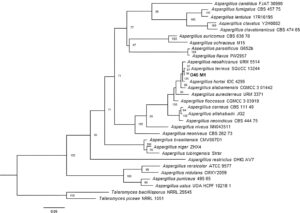
Pada pohon filogenetik yang dibangun untuk isolat genus Aspergillus , O45M1 (Gambar 2 ), juga diamati bahwa anggota bagian-bagian berbeda dari genus tersebut diorganisasikan ke dalam klade yang berbeda dan penyelarasan sekuens isolat O45M1 dalam klade Terrei, khususnya, sesuai dengan sekuens lain dari spesies A. terreus .
3.2 Kecernaan in vitro serat bahan kering dan serat deterjen netral jerami Urochloa decumbens
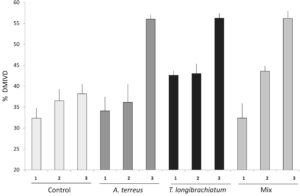
DMIVD jerami U. decumbens dipengaruhi secara signifikan oleh keberadaan T. longibrachiatum ( p < .01) dan oleh periode pengumpulan cairan rumen ( p < .01). Inokulasi T. longibrachiatum mendorong tingkat DMIVD yang lebih tinggi untuk tiga periode pengumpulan cairan rumen (Gambar 3 , p < .01). Namun, DMIVD lebih rendah pada periode pengumpulan pertama dibandingkan dengan yang diamati pada periode kedua dan ketiga ( p < .01).
Tingkat NDFD pada periode ketiga secara signifikan lebih rendah daripada yang diamati pada periode pertama dan kedua, untuk kedua inokulum (Tabel 2 , p < .01, dengan uji Fisher LSD). Keberadaan inokulum ini tidak memengaruhi NDFD ( p > .05).
| Periode | Kontrol | Mobil B13 M2 | O45 M1 | Mencampur | Rata-rata periode | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 41.58 | 0,91 | 41.83 | 2.61 | 41.61 | 0,70 | 42.59 | 0,97 | 42.24 | 2.16 |
| 2 | 42.43 | 0,59 | Tanggal 41.09 | 0,93 | 41.45 | 2.88 | 41.99 | 0.45 | 41.65 | 2.08 |
| 3 | 37.28 | 0,88 | 39.26 | 0,70 | 38.56 | 0,90 | 38.37 | 1.07 | 38.37 | 1.76 |
Catatan : B13 M2 isolat – Aspergillus terreus ; O45 M1 isolat – Trichoderma longibrachiatum ; Campuran = campuran dua jamur dan kontrol dievaluasi dalam periode pengumpulan cairan rumen. Rata-rata antara inokulan secara statistik serupa ( p > .05). Rata-rata pada periode ketiga lebih rendah daripada periode pertama dan kedua (LSD untuk periode = 1,852 derajat kebebasan residual = 84, p < .01).
3.3 Kuantifikasi jamur selama analisis kecernaan
Kehadiran jamur diverifikasi dalam kultur dari pengumpulan cairan rumen untuk sampel yang dievaluasi pada awal inkubasi. Inokulasi T. longibrachiatum ( p <.01) dan waktu inkubasi ( p <.01) secara signifikan memengaruhi penghitungan jamur ini dalam medium yang mengandung cairan rumen (Tabel 3 ). Untuk sampel yang dikumpulkan pada 72 jam inkubasi (setelah pencernaan asam), populasi jamur miselium lebih tinggi ketika T. longibrachiatum diinokulasi daripada yang diamati untuk inokula lain (Tabel 3 , p <.01). Namun, konsentrasi rata-rata jamur berkurang pada 72 jam inkubasi untuk kedua inokulan yang dievaluasi ( p <.01 dengan LSD) dengan membandingkan 0 dan 24 jam inkubasi (Tabel 3 ).
| Inokulum | 0 jam | SEJARAH | 24 jam | SEJARAH | 72 jam | SEJARAH | Rata-rata inokulum* |
|---|---|---|---|---|---|---|---|
| Trichoderma longibrachiatum | 1,2 × 10 4 | 17 | 7,0 × 10 3 | 98 | 3,3 × 10 3 | 55 | 3.77 |
| Aspergillus terreus | 7,0 × 10 3 | 10 | 3,0 × 10 3 | 94 | 2,0 × 10 1 | angka 0 | 3.04 |
| Campuran jamur | 2,3 × 10 3 | 23 | 6,0 × 10 3 | 30 | 1,0 × 10 1 | 11 | 3.35 |
| Kontrol | 2,3 × 10 3 | 26 | 3,0 × 10 3 | 50 | 1,0 × 10 1 | angka 0 | 2.77 |
| Rata-rata periode* | 3.73 | 0.28 | 3.75
0.733 |
2.16
0,96 |
Catatan : Interaksi inokulasi T. longibrachiatum dan A. terreus × periode inkubasi signifikan ( p = .0217, LSD = 0,643). Konsentrasi jamur pada 72 jam inkubasi lebih tinggi ketika T. loginbrachiatum diisolasi ( p < .01). Setelah pencernaan asam pada 72 jam inkubasi, rata-rata konsentrasi jamur lebih rendah dari nol dan 24 jam ( p < .01).
4 DISKUSI
4.1 Karakterisasi molekular isolat jamur
Spesies T. longibrachiatum dan A. terreus yang diisolasi dari usus besar domba yang merumput di padang rumput tropis juga telah terdeteksi dalam cairan rumen sapi dari berbagai usia. Studi tentang jamur miselium sapi zebu yang dibesarkan di padang rumput tropis di wilayah semi-kering mengidentifikasi A. terreus pada anak sapi, sapi jantan dan sapi betina, sementara T. longibrachiatum diisolasi dalam sampel cairan rumen dari sapi jantan (Abrão et al., 2014 ). Dalam studi lain, mikrobiota aerobik dari cairan rumen dari sapi Holstein dan sapi dara yang diberi makan hijauan tropis yang berbeda dianalisis, dan genus Aspergillus adalah yang paling sering diamati, sesuai dengan 56% dari isolat. Namun, genus Aspergillus , Gliocladium , Paecilomyces , Rhizopus , Scedosporium dan Trichoderma juga diidentifikasi (Almeida et al., 2014 ).
Karakterisasi isolat A. terreus dari rumen sapi menunjukkan aktivitas avicelase dan xilanase yang lebih tinggi dibandingkan spesies Aspergillus lainnya (Abrão et al., 2017 ). Penulis melaporkan bahwa isolat terpilih menghasilkan enzim tingkat tinggi untuk degradasi dinding sel vegetal, yang menunjukkan potensi untuk digunakan sebagai probiotik atau aditif mikroba untuk diet ruminansia.
4.2 Kecernaan in vitro serat bahan kering dan serat deterjen netral jerami Urochloa decumbens
DMIVD jerami U. decumbens yang dievaluasi dalam percobaan ini berkisar antara 32,0% hingga 56,0%. Pakan ini memiliki kualitas nutrisi yang rendah, menunjukkan kadar NDF, ADF, dan lignin yang tinggi serta kadar protein kasar, ekstrak ethereal, dan gula total yang rendah. Pakan ini dikumpulkan setelah berbunga, pada awal musim kemarau, yang membenarkan rendahnya daya cerna yang disebabkan oleh komposisi nutrisi yang buruk.
Peningkatan kecernaan DM dari jerami lignifikasi U. decumbens dengan penyertaan T. longibrachiatum dapat dikaitkan dengan degradasi fraksi berserat dari hijauan ini yang disebabkan oleh aksi hidrolitik selulase yang diproduksi oleh jamur ini, yang menyediakan karbohidrat yang mudah dicerna oleh mikroorganisme cairan rumen. Jamur dari genus Trichoderma dapat menghasilkan p -glukosidase (Li et al., 2016 ), xilanase (Toth et al., 2013 ), karboksimetilselulase, avicellase, β-glukosidase, β-xilosidase, α-arabinofuranosidase, β-galaktosidase (Morgavi et al., 2000 ) dan hemiselulase (Schuster & Schmoll, 2010 ).
Tingkat IVDMD tertinggi diamati ketika isolat T. longibrachiatum diinokulasi dalam cairan rumen yang dikumpulkan pada periode kedua dan ketiga. Domba yang menyumbangkan cairan rumen mulai menerima diet dengan proporsi jerami UD yang lebih tinggi pada fase adaptasi percobaan. Mungkin, mikrobiota rumen domba yang berfistula beradaptasi lebih baik dalam dua periode terakhir ini, dan penambahan jamur ini berkontribusi pada IVDMD yang lebih baik bersama dengan mikroorganisme rumen hewan.
Dalam penelitian ini, ekspresi enzim yang lebih besar oleh T. longibrachiatum akan berkontribusi pada degradasi dinding sel U. decumbens yang lebih baik dibandingkan dengan isolat A. terreus . Dengan menggunakan biomassa sorgum, Nurudeen et al. ( 2015 ) memverifikasi bahwa isolat T. longibrachiatum dan A. terreus masing-masing menghasilkan 1,82 dan 0,95 U/mL karboksimetil selulosa.
Hasil-hasil ini menunjukkan potensi penggunaan jamur ini sebagai aditif mikroba dalam diet ruminansia yang diberi pakan hijauan tropis dan menguatkan penelitian lain yang mengevaluasi penyertaan enzim jamur miselium untuk meningkatkan daya cerna hijauan (Nurudeen et al., 2015 ). Selain itu, satu penelitian menunjukkan bahwa penyertaan enzim fibrolitik dari T. longibrachiatum dalam diet sapi perah meningkatkan jumlah bakteri rumen yang memanfaatkan hemiselulosa dan produk sekunder dari pencernaan selulosa (Nsereko et al., 2002 ).
Dalam studi lain, efek penambahan selulase Aspergillus niger , T. longibrachiatum dan campuran selulase dari jamur ini dievaluasi pada kecernaan in vitro dari diet yang mengandung 70% jerami rumput dan 30% konsentrat. Penambahan selulase eksogen ini meningkatkan DMIVD dan NDFD yang lebih tinggi setelah 24 jam inkubasi; namun, pada 48 jam, efek ini tidak diamati, yang menunjukkan bahwa penambahan selulase eksogen hanya merangsang fase awal degradasi (Giraldo et al., 2007 ). Secara berbeda, dalam studi saat ini, ketika T. longibrachiatum digunakan sebagai inokulum, ia meningkatkan peningkatan kecernaan ini bahkan setelah 48 jam inkubasi, yang menunjukkan produksi selulase oleh T. longibrachiatum selama fermentasi in vitro dalam cairan rumen.
Berbagai jenis rumput yang digunakan di padang rumput dievaluasi sebagai substrat untuk enzim fibrolitik Aspergillus japonicus CO3 dalam fermentasi terendam (Facchini et al., 2011 ). Padang rumput dari genus Urochloa adalah yang menunjukkan tingkat degradasi yang lebih baik dengan ekstrak enzimatik yang disertakan. Menurut penulis, meskipun hijauan tropis memiliki nilai gizi yang rendah, penggunaan ekstrak enzimatik sebagai praperlakuan makanan ini dapat berkontribusi pada degradasi serat yang lebih baik, yang mendukung mikrobiota rumen.
Dalam penelitian ini, untuk periode ketiga pengumpulan cairan rumen, tingkat NDFD lebih rendah daripada yang diamati pada periode pengumpulan pertama dan kedua. Mikrobiota rumen hewan-hewan ini mungkin telah beradaptasi lebih baik pada periode ketiga untuk degradasi ADF dan komponen berserat lainnya dari hijauan ini dengan lebih baik. Dengan cara ini, akan ada cadangan enzimatik yang lebih rendah untuk mendegradasi komponen NDF, yang harus dievaluasi dalam penelitian selanjutnya. Penelitian lebih lanjut, dengan hewan yang tidak beradaptasi dengan diet tinggi serat, juga harus dilakukan dengan penambahan jamur ini untuk mengevaluasi potensi bioteknologi sebagai aditif atau probiotik untuk ruminansia dalam fase transisi ke diet tinggi serat pada periode kering dan untuk diet pasca-sapih.
4.3 Kuantifikasi jamur selama analisis kecernaan
Jumlah jamur miselium diamati dalam cairan rumen tanpa inokulasi strain yang dipilih, yang menunjukkan bahwa kelompok jamur ini secara alami hadir dalam cairan rumen domba, seperti yang ditunjukkan dalam penelitian sebelumnya pada hewan yang diberi makan di padang rumput tropis (Freitas et al., 2012 ). Pertumbuhan yang diamati untuk T. longibrachiatum setelah 72 jam dalam kondisi abomasum yang disimulasikan sesuai dengan karakterisasi genus Trichoderma , yang tumbuh dalam kisaran pH 4 dan 5, dengan hanya beberapa spesies yang mendukung nilai pH <3 (Chahal et al., 1992 ). Mikroorganisme dengan potensi probiotik harus menunjukkan resistensi dalam kondisi keasaman lambung yang disimulasikan (Angelakis, 2017 ). Hasil yang diamati dalam penelitian ini dapat menunjukkan potensi bioteknologi dari isolat T. longibrachiatum untuk digunakan dalam formulasi probiotik komersial baru untuk ruminansia.
5 KESIMPULAN
Analisis sekuens dari kedua wilayah ITS rDNA dan fragmen gen β-tubulin yang mengkode diperlukan untuk identifikasi akurat isolat O45M1 sebagai Aspergillus terreus . Namun, isolat B13M2 dapat diidentifikasi sebagai Trichoderma longibrachiatum , dengan mempertimbangkan analisis sekuens wilayah ITS dan gen yang mengkode faktor pemanjangan 1-α. Penambahan isolat T. longibrachiatum mendorong peningkatan signifikan dalam DMIVD untuk jerami lignifikasi UD, yang menunjukkan potensi yang menjanjikan untuk pengembangan aditif probiotik atau mikroba untuk penggembalaan domba di padang rumput dengan nilai gizi rendah.