
Abstrak
Intensifikasi pertanian merupakan penyebab utama hilangnya keanekaragaman hayati di seluruh dunia. Akan tetapi, agroekosistem tradisional sering dikaitkan dengan keanekaragaman burung yang tinggi karena heterogenitas bentang alamnya, yang menawarkan relung yang tersedia bagi berbagai spesies burung. Di sini, kami berfokus pada perubahan temporal dalam keanekaragaman taksonomi, fungsional, dan filogenetik komunitas burung dari bentang alam pertanian tradisional Satoyama di Jepang. Kami menemukan tren temporal yang signifikan (misalnya peningkatan) dalam kekayaan spesies secara keseluruhan, kekayaan spesies spesialis hutan, keanekaragaman filogenetik, dan keterkaitan filogenetik dalam kelompok burung, terlepas dari komposisi penggunaan lahan di sekitar lokasi tersebut. Peningkatan kekayaan spesies dan keterkaitan filogenetik secara simultan dapat menyoroti proses homogenisasi biotik, yang merupakan ciri khas lingkungan yang mengalami antropisasi. Keanekaragaman burung juga dipengaruhi secara signifikan oleh proporsi badan air (misalnya peningkatan kekayaan dan penyebaran fungsional, tetapi penurunan keseragaman atau redundansi fungsional) dan jenis penggunaan lahan lainnya (misalnya hubungan negatif antara kekayaan spesies dan proporsi ladang). Proporsi sawah memengaruhi setiap jenis kekayaan burung secara berbeda: bentuk U terbalik untuk generalis hutan, negatif untuk spesies spesialis hutan, dan positif untuk spesialis lahan terbuka. Ketika menilai stabilitas temporal komposisi komunitas burung, kami menemukan bahwa stabilitas tersebut berkorelasi secara signifikan dengan proporsi padang rumput, badan air, dan lanskap perkotaan. Secara khusus, komunitas burung yang dikelilingi oleh padang rumput dicirikan oleh penggantian spesies yang lebih tinggi dari waktu ke waktu. Selain itu, proporsi lanskap perkotaan yang sangat rendah atau sangat tinggi dikaitkan dengan ketidakstabilan relatif komposisi komunitas burung. Temuan kami mendukung hipotesis bahwa sistem pertanian tradisional mewakili lanskap berharga yang mendukung keanekaragaman burung. Namun, komposisi relatif jenis penggunaan lahan sangat penting dalam membentuk berbagai komponen keanekaragaman taksonomi, fungsional, dan filogenetik dalam kumpulan burung dan stabilitas temporalnya.
Perkenalan
Bentang alam pertanian adalah jenis lingkungan antropogenik terbesar dalam skala global (Foley et al. 2011 ). Jenis bentang alam seperti itu juga merupakan habitat bagi banyak spesies satwa liar, yang memelihara beberapa komunitas, terutama bagi burung-burung di lahan pertanian (Wuczyński et al. 2011 , Herzon et al. 2014 , Sasaki et al. 2020 ). Secara keseluruhan, keanekaragaman hayati telah beradaptasi dan dibentuk oleh sejarah dan perubahan penggunaan lahan pertanian (Emmerson et al. 2016 ). Komposisi dan pengaturan spasial jenis penggunaan lahan sangat berbeda di antara agroekosistem (Sasaki et al. 2020 ). Bentang alam pertanian secara kasar dapat menggambarkan gradien agroekosistem yang besar, dari daerah yang homogen dan heterogen hingga pengelolaan pertanian yang ekstensif atau intensif. Diversifikasi agroekosistem yang luas dan dampak penggunaan lahan di sekitarnya adalah faktor kunci yang menjelaskan perubahan utama dalam keanekaragaman satwa liar setempat (Kleijn et al. 2009 , Santana et al. 2017 ). Telah diketahui dengan baik bahwa intensifikasi pertanian (misalnya mekanisasi, penggunaan agrokimia, penggantian polikultur dengan monokultur) telah mengurangi heterogenitas habitat (Bignal dan McCracken 2000 , Benton et al. 2003 , Santana et al. 2017 ), mengurangi ketersediaan habitat berkualitas untuk spesies satwa liar (Kleijn et al. 2009 , Tscharntke et al. 2012 , Morelli et al. 2017a ), dan juga berdampak pada fungsi ekosistem (Emmerson et al. 2016 ).
Di Eropa, praktik pertanian intensif mendorong penurunan dramatis dalam populasi burung (Rigal et al. 2023 ). Namun, lanskap pertanian tradisional dapat dikaitkan dengan konservasi keseluruhan keanekaragaman hayati pada skala spasial yang berbeda, terutama mengenai keanekaragaman burung (Morelli et al. 2014 , Lomba et al. 2015 , Maskell et al. 2019 ). Heterogenitas spasial lanskap dianggap sebagai faktor kunci untuk distribusi keanekaragaman hayati secara keseluruhan di daerah pertanian tradisional (Benton et al. 2003 , Fahrig et al. 2011 ). Kemunculan beberapa spesies burung sangat terkait dengan jumlah vegetasi marginal yang tidak produktif di lahan pertanian (Ceresa et al. 2012 , Morelli 2013 , Regos et al. 2016 ). Selain itu, dalam agroekosistem Jepang, ditemukan bahwa bentang alam heterogen mendukung komunitas burung yang lebih kaya, terutama spesies yang memiliki jangkauan luas (Katayama et al. 2014 ). Bentang alam Satoyama adalah contoh yang baik dari bentang alam pertanian tradisional yang membantu menjaga keanekaragaman hayati lokal (Katoh et al. 2009 ) dan juga dianggap sebagai sistem yang berkelanjutan (Washitani 2001 ). Nama jenis bentang alam ini terdiri dari ‘Sato’, yang berarti pemukiman, dan ‘yama’, yang berarti gunung, bukit, atau hutan (Katoh et al. 2009 ). Lebih tepatnya, bentang alam Satoyama adalah mosaik kompleks yang dibentuk oleh beberapa habitat semi-alami, yang meningkatkan heterogenitas bentang alam secara keseluruhan (Katoh et al. 2009 ).
Beberapa karakteristik khusus penggunaan lahan juga dapat membentuk komposisi dan kekayaan spesies burung di komunitas lahan pertanian (Faria et al. 2024 ). Namun, kekayaan spesies tidak selalu menjadi alat terbaik untuk berfokus pada keanekaragaman burung. Bahkan jika itu merupakan metrik yang lebih sederhana untuk mengukur keanekaragaman dalam komunitas burung (Magurran 2004 ), alat ini mungkin tidak memadai karena mengabaikan identitas spesies, peran fungsional, dan kelimpahan relatif. Akibatnya, pendekatan ini dihukum karena kurangnya pertimbangan mengenai peran ekologis spesies dalam kumpulan (Safi et al. 2013 ). Metrik keanekaragaman fungsional, misalnya, lebih cocok untuk menjelaskan perubahan dalam fungsi ekosistem daripada metrik keanekaragaman taksonomi, karena mereka berfokus pada peran fungsional s (Petchey et al. 2004 ). Ukuran redundansi fungsional dan penyebaran fungsional dari himpunan spesies dapat dikaitkan dengan tingkat ketahanan ekosistem, yang berpotensi meningkatkan kapasitasnya untuk menyerap tekanan lingkungan (Gerisch et al. 2012 , Lee and Martin 2017 ). Redundansi fungsional yang tinggi di antara komponen-komponen suatu sistem dapat meningkatkan elastisitas sistem secara keseluruhan (Haegeman et al. 2016 ). Ini karena spesies dengan sifat yang serupa (misalnya redundansi tinggi, penyebaran fungsional lebih rendah) memainkan peran yang analog dalam proses ekosistem (Bello et al. 2007 , Laureto et al. 2015 ). Metrik keanekaragaman filogenetik memberikan informasi pelengkap untuk menggambarkan keanekaragaman hayati, yang terkait dengan warisan evolusi (Laity et al. 2015 ). Sejarah evolusi adalah bagian penting yang menjelaskan himpunan spesies di lingkungan yang berbeda. Misalnya, spesies dari klade yang baru-baru ini terdiversifikasi cenderung menjadi ‘pemanfaat’ habitat pertanian yang lebih baik, yang berpotensi mengambil keuntungan dari intensifikasi pertanian (Frishkoff et al. 2014 ). Selain itu, beberapa metrik filogenetik yang dihitung pada komunitas dapat dikaitkan dengan ‘respons ekologis’ potensial mereka saat menghadapi stresor. Keterkaitan filogenetik, misalnya, menggambarkan tingkat asosiasi filogenetik di antara spesies dalam suatu himpunan. Singkatnya, dua spesies lebih berkorelasi secara filogenetik jika mereka memiliki nenek moyang yang lebih baru (Godoy et al. 2014 ). Metrik filogenetik ini, sebagai tambahan, diharapkan dikaitkan dengan koeksistensi spesies dan aturan kompetitif (Losos 2008 , Godoy et al. 2014 ).
Dampak karakteristik khusus lanskap pertanian tradisional Jepang ini terhadap keanekaragaman burung belum diukur dalam kerangka kerja multi-segi. Kami perkirakan bahwa setiap komponen berbeda yang menggambarkan keanekaragaman burung di habitat pertanian (misalnya taksonomi, fungsional, atau filogenetik) dapat dipengaruhi secara berbeda oleh stresor lingkungan (Lee dan Martin 2017 , Morelli et al. 2017b ). Modifikasi semacam itu seharusnya lebih diperhatikan saat berfokus pada perubahan lintas garis waktu, seperti yang disorot dalam studi sebelumnya (Harrison et al. 2014 , Chalmandrier et al. 2015 , Costa et al. 2020 , Floigl et al. 2022 ). Akibatnya, studi ini mengeksplorasi perubahan temporal dalam keanekaragaman taksonomi (kekayaan spesies), keanekaragaman fungsional, dan keanekaragaman filogenetik komunitas burung selama sepuluh tahun di lanskap Satoyama di Jepang. Kami menguji karakteristik komposisi penggunaan lahan mana yang memengaruhi dimensi keanekaragaman burung di salah satu negara dengan populasi paling rendah di dunia, dan juga mengeksplorasi bagaimana berbagai jenis penggunaan lahan dapat membentuk stabilitas temporal komposisi komunitas burung.
Bahan dan metode
Komunitas burung dan metrik keanekaragaman
Studi ini menggunakan data burung dari Proyek ‘Monitoring Sites 1000’, survei pemantauan keanekaragaman hayati di seluruh ekosistem darat dan perairan Jepang (Pusat Keanekaragaman Hayati Jepang 2022 ). Kami berfokus pada kumpulan data yang terdiri dari 106 lokasi yang dicirikan oleh lanskap Satoyama, yang dikunjungi selama sepuluh tahun (2008‒2017). Lokasi-lokasi ini dipilih secara tidak acak, tetapi upaya survei didistribusikan secara relatif merata (Katayama et al. 2024 ). Setiap lokasi dikunjungi hingga empat kali selama musim kawin (April hingga Juli), dari pukul 04:00 hingga 09:00 hanya pada hari-hari tanpa hujan atau angin kencang, untuk meminimalkan variasi dalam detektabilitas (Katayama et al. 2014 ). Semua individu burung yang terdeteksi dalam radius 50 m dicatat (Kawamura et al. 2019 , Katayama et al. 2024 ), dan analisis lebih lanjut didasarkan pada kemunculan setiap spesies burung (misalnya data ada-tidaknya).
Dengan menggunakan matriks kemunculan spesies burung di seluruh lokasi survei, berbagai metrik komunitas dihitung untuk mengeksplorasi tiga dimensi utama:
1) Keanekaragaman taksonomi (jumlah seluruh spesies dalam suatu himpunan atau kekayaan spesies total, dan kekayaan spesies yang diuraikan berdasarkan kelompok habitat dan spesialisasi sebagai spesialis hutan, generalis hutan, spesialis lahan terbuka dan kekayaan spesies spesies lain (Katayama et al. 2024 ),
2) keragaman fungsional (FRic – kekayaan fungsional, FEve – keseragaman fungsional, FDis – dispersi fungsional dan RaoQ – entropi kuadrat Rao), dan
3) keanekaragaman filogenetik (PD dan PSV – keterkaitan filogenetik).
Metrik keragaman fungsional yang diterapkan dalam studi ini saling melengkapi dan menggambarkan berbagai komponen yang terkait dengan peran fungsional spesies dalam perakitan. Ciri-ciri fungsional burung diekstraksi dari Wilman et al. ( 2014 ), berdasarkan ekologi makan, aktivitas harian, dan massa tubuh. Dua puluh variabel menggambarkan relung setiap spesies, dalam hal 1) jenis diet (10 variabel), 2) substrat tempat makanan diambil (8 variabel), 3) periode hari saat spesies mencari makan secara aktif (1 variabel), dan 4) massa tubuh dalam gram (1 variabel). Semua variabel mengenai jenis diet dan substrat mencari makan dinyatakan sebagai persentase (berkisar dari 0 hingga 100) (Wilman et al. 2014 ), dan jenis sifat Eltonian tersebut digunakan secara luas dalam studi sebelumnya untuk mengkarakterisasi komunitas burung (Benedetti et al. 2005 , Morelli et al. 2021 , Zbyryt et al. 2021 , Shuai et al. 2024 ). Kekayaan fungsional adalah jumlah ruang fungsional yang ditempati oleh suatu himpunan spesies (Schleuter et al. 2010 ). Keseragaman fungsional menggambarkan keteraturan distribusi biomassa dalam himpunan tersebut mengenai seluruh ruang relung, yang menjadi ukuran redundansi fungsional (Villéger et al. 2008 ). Dispersi fungsional mengukur seberapa luas ciri-ciri fungsional atau peran ekologis di seluruh himpunan spesies, dengan dispersi yang lebih tinggi dikaitkan dengan rentang respons yang lebih luas terhadap potensi stres lingkungan (Laliberté et al. 2010 , Cooke et al. 2019 ). Akhirnya, entropi kuadrat Rao didasarkan pada jumlah jarak berpasangan antara spesies mengenai berbagai ciri mereka (Zoltan 2005 ). Semua indeks keragaman fungsional yang disebutkan di atas dihitung menggunakan paket ‘FD’ untuk R (Laliberté et al. 2015 ).
PD menunjukkan jumlah sejarah evolusi dalam suatu himpunan spesies, dihitung dengan mempertimbangkan panjangnya jalur bersama pada filogeni yang direpresentasikan oleh sekumpulan taksa (Faith 1992 , Faith dan Baker 2007 , Tucker et al. 2016 ). PSV mengukur tingkat keterkaitan filogenetik semua spesies burung dalam himpunan tersebut (Helmus et al. 2007 ). Untuk memperkirakan PD dan PSV, kami membangun pohon filogenetik menggunakan hubungan di antara spesies pada setiap hitungan titik, berdasarkan data genetik semua spesies burung yang tercatat dalam penelitian ini (Jetz et al. 2012 ) yang diunduh dari ‘BirdTree’ ( www.birdtree.org ) dan mempertimbangkan pohon konsensus yang diperoleh dengan fungsi ‘konsensus’ pada 100 pohon acak, dengan paket ‘ape’ ver. 5.3 untuk R (Paradis et al. 2004 ). PD dan PSV dihitung menggunakan ‘Picante’ ver. 1.7 paket untuk R (Kembel dkk. 2010 ).
Karakteristik penutup lahan
Bahasa Indonesia: Di sekitar setiap lokasi pengambilan sampel (misalnya jumlah titik), menggunakan buffer 1 km, kami menghitung komposisi penggunaan lahan. Buffer 1 km dipilih karena ukuran tersebut merupakan kompromi yang baik antara luas dan akurasi untuk mengkarakterisasi komposisi area sekitar pada skala spasial yang besar (misalnya nasional) dan telah dieksplorasi dalam studi sebelumnya untuk melakukan model distribusi spesies untuk spesies burung (Clavero et al. 2011 , Villaseñor dan Escobar 2019 ). Data penggunaan lahan diekstraksi dari Peta Digital Survei Vegetasi skala 1:25.000 dalam GIS Informasi Lingkungan Alam yang dikembangkan oleh Kementerian Lingkungan Hidup ( http://gis.biodic.go.jp/webgis/sc-023.html ). Kemudian, untuk menghindari informasi yang berlebihan, kami menggabungkan beberapa kelas penggunaan lahan untuk mendapatkan deskripsi yang akurat dari lingkungan sekitar, yang dijelaskan dengan tujuh kategori penggunaan lahan berikut: Padang rumput, hutan, air, ladang (atau lahan pertanian), sawah, perkotaan, dan jenis penggunaan lahan lainnya. Dengan menggunakan operator ‘intersect’ dari QGIS (Quantum GIS Development Team 2024 ), kami memperkirakan luas untuk setiap kategori penggunaan lahan. Akhirnya, area cakupan diubah menjadi proporsi dan digunakan sebagai deskriptor untuk setiap lokasi pengambilan sampel tunggal.
Analisis data
Kami telah menggunakan model campuran aditif umum (GAMM) (Wood 2017 ) untuk mengeksplorasi tren temporal dalam keanekaragaman taksonomi (kekayaan spesies total dan kekayaan gugus fungsi spesifik), keanekaragaman fungsional (FRic, FEve, FDis dan RaoQ) dan keanekaragaman filogenetik (PD dan PSV) komunitas burung yang diamati antara tahun 2008 dan 2017 di 106 lokasi. Pemodelan aditif adalah pendekatan semi-parametrik fleksibel yang memungkinkan kami untuk mengeksplorasi berbagai pola, termasuk tren non-linier sambil menyeimbangkan kecocokan model dan kompleksitasnya (Hastie dan Tibshirani 1986 ). Perubahan temporal direpresentasikan menggunakan spline regresi pelat tipis (Wood 2003 ) waktu dengan dimensi dasar peringkat lima. Mengingat bahwa lokasi tersebut diamati selama beberapa tahun, kami mengatasi masalah pengamatan berulang untuk lokasi yang sama dengan menggabungkan intersepsi acak untuk lokasi di GAMM. Nilai non-negatif dari karakteristik keanekaragaman dimodelkan sebagai respons Gaussian menggunakan fungsi tautan logaritmik. Data jumlah kekayaan spesies disesuaikan menggunakan model Poisson. Karena beberapa model kekayaan menunjukkan varians yang lebih rendah daripada yang diasumsikan oleh distribusi kesalahan Poisson, data disesuaikan kembali menggunakan pendekatan quasi-likelihood untuk memperhitungkan underdispersion (Wood 2017 ).
Karena pola temporal dalam keanekaragaman burung dapat dipengaruhi oleh perubahan dalam penggunaan lahan di sekitarnya, kami menerapkan serangkaian GAMM yang, di samping efek waktu yang tetap, menggabungkan berbagai karakteristik penggunaan lahan, termasuk proporsi padang rumput, perairan, ladang, sawah, dan lanskap perkotaan. Tutupan hutan dan substrat lainnya tidak terlibat dalam model karena korelasi yang kuat dengan tutupan ladang (r = ‒0,7) dan proporsi keseluruhan yang rendah (tutupan maks. < 10%). Kami menggunakan parameterisasi model yang sama seperti di atas, tetapi menerapkan penalti ganda untuk memfasilitasi penyusutan istilah yang tidak informatif menuju nol, sehingga memungkinkan pemilihan variabel otomatis (Marra dan Wood 2011 ).
Akhirnya, kami mengeksplorasi stabilitas temporal komunitas burung dan pengaruh perubahan tutupan lahan dalam proses ini. Untuk mengukur stabilitas temporal komposisi komunitas, kami menggunakan matriks kelimpahan waktu-demi-spesies dan menghitung ketidakmiripan komunitas menggunakan jarak Bray–Curtis (Bray dan Curtis 1957 ) untuk setiap lokasi. Komunitas yang stabil dengan pergantian spesies yang rendah dan kelimpahan yang serupa diharapkan menunjukkan kesamaan komposisi komunitas yang tinggi, sementara komunitas yang kurang stabil yang mengalami pertukaran spesies yang lebih tinggi dan kelimpahan yang berfluktuasi diharapkan menunjukkan kesamaan yang rendah dari waktu ke waktu. Di setiap lokasi, kami membagi ketidakmiripan keseluruhan menjadi dua komponen 1) variasi seimbang dalam kelimpahan, di mana individu dari beberapa spesies digantikan oleh jumlah individu yang sama dari spesies yang berbeda dari waktu ke waktu, dan 2) gradien kelimpahan, di mana beberapa individu hilang dari satu tahun ke tahun lainnya (Baselga 2017 ). Komponen variasi seimbang dan gradien kelimpahan dianalisis dalam kerangka model aditif. Namun, tidak ada efek acak yang ditentukan, karena rangkaian waktu dipecah menjadi dua komponen per situs. Karena komponen ketidaksamaan dibatasi antara nol (menunjukkan stabilitas absolut dalam waktu) dan satu, kami memodelkan komponen ini sebagai respons beta menggunakan fungsi tautan logit (Ferrari dan Cribari-Neto 2004 ).
Fungsi autokorelasi dan korelogram spline (Bjørnstad dan Falck 2001 ) digunakan untuk memeriksa pola autokorelasi temporal dan spasial dalam residual semua model. Selain itu, plot diagnostik standar dari residual dibuat untuk mengevaluasi asumsi distribusional. Tidak ada autokorelasi substansial atau penyimpangan dari distribusi yang diasumsikan yang diamati. Signifikansi istilah model dinilai menggunakan uji tipe Wald (Wood 2013a ). Kesesuaian model secara keseluruhan dievaluasi menggunakan koefisien determinasi yang disesuaikan. Analisis dilakukan dalam R ver. 4.2.2 ( www.r-project.org ) menggunakan pustaka ‘betapart’ (Baselga et al. 2023 ), ‘ggplot2’ (Wickham 2016 ), ‘ncf’ (Bjørnstad 2020 ) dan ‘mgcv’ (Wood 2013b ).
Hasil
Sebanyak 179 spesies burung tercatat di 106 lokasi yang dicirikan oleh lanskap pertanian tradisional Satoyama selama 10 tahun di Jepang (Informasi pendukung). Spesies burung yang paling sering terlihat di lanskap pertanian adalah burung bulbul bertelinga cokelat Hypsipetes amaurotis , burung warbler semak Jepang Horornis diphone , burung tit Asia Parus cinereus dan burung gagak berparuh besar Corvus macrorhynchos , yang hadir di lebih dari 90% catatan (Informasi pendukung).
Nilai keanekaragaman taksonomi, fungsi, dan filogenetik pada komunitas burung di lanskap Satoyama menunjukkan pola yang berbeda selama 10 tahun (Informasi pendukung). Pembuatan model campuran aditif menunjukkan tren temporal yang signifikan dalam kekayaan spesies total (derajat kebebasan efektif [edf] = 1,0, F = 6,5, p = 0,011, R 2 = 0,635), kekayaan spesies spesialis hutan (edf = 1,0, F = 15,1, p < 0,001, R 2 = 0,832), FEve (edf = 1,0, F = 4,1, p = 0,042, R 2 = 0,346), PD (edf = 3,1, F = 4,4, p = 0,002, R 2 = 0,553) dan PSV (edf = 1,9, F = 6,1, p = 0,002, R 2 = 0,553). Sebaliknya, pola kekayaan spesies generalis hutan (edf = 1.0, F = 0.1, p = 0.739, R 2 = 0.572), spesialis lahan terbuka (edf = 1.0, F = 0.1, p = 0.774, R 2 = 0.774), kekayaan spesies lain (edf = 2.0, F = 3.6, p = 0.195, R 2 = 0.398), FRic (edf = 1.0, F = 0.8, p = 0.361, R 2 = 0.469), FDis (edf = 1.0, F = 0.1, p = 0.771, R 2 = 0.486), dan RaoQ (edf = 1.0, F < 0.1, p = 0.889, R 2 = 0,491) tidak signifikan. Semua model signifikan menunjukkan peningkatan yang stabil dalam keanekaragaman komunitas burung dari waktu ke waktu (Gbr. 1 , Informasi pendukung).

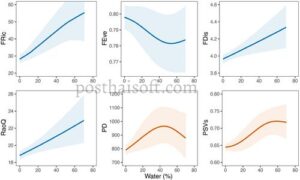
Ketika kami memasukkan karakteristik penutup lahan ke dalam GAMM, semua pola keragaman yang awalnya diidentifikasi meningkat dari waktu ke waktu tetap signifikan kecuali untuk FEve, yang menjadi sedikit tidak signifikan (Tabel 1 ). Di antara karakteristik penutup lahan, proporsi badan air secara signifikan terkait dengan perubahan pada sebagian besar indeks keragaman. FRic, FDis dan RaoQ meningkat dengan proporsi badan air. Pada saat yang sama, FEve menurun, dan indeks keragaman filogenetik menunjukkan pola unimodal (Gbr. 2 ). Karakteristik penutup lahan lainnya juga secara signifikan memengaruhi indeks keragaman burung (Tabel 1 ). Indeks keragaman fungsional FEve dan FDis dipengaruhi secara negatif oleh proporsi padang rumput (Gbr. 3 ). Sebaliknya, FRic, FDis dan RaoQ secara positif terkait dengan proporsi lanskap perkotaan. Hubungan positif juga diamati antara proporsi sawah dan FRic dan PSV. Kekayaan spesies burung total secara signifikan dipengaruhi secara negatif oleh proporsi ladang (Tabel 1 , Gbr. 3 ). Peningkatan proporsi tipe tutupan lahan non-hutan berkorelasi negatif dengan kekayaan spesies spesialis hutan. Sebaliknya, spesialis lahan terbuka dan spesies lain berkorelasi positif dengan tipe tutupan lahan ini (Tabel 1 , Informasi pendukung). Terakhir, kekayaan spesies generalis hutan menunjukkan respons unimodal terhadap proporsi sawah di lanskap (Tabel 1 , Informasi pendukung).

Tabel 1. Ringkasan model aditif yang menguji pengaruh waktu dan perubahan jenis tutupan lahan terhadap keanekaragaman taksonomi, keanekaragaman fungsional, keanekaragaman filogenetik, dan stabilitas temporal komunitas burung di lanskap pertanian Satoyama, Jepang, selama sepuluh tahun. Derajat kebebasan efektif (edf), statistik uji (F dalam model keanekaragaman dan χ 2 dalam model stabilitas), dan probabilitas terkait (p) ditampilkan untuk istilah model. Koefisien determinasi yang disesuaikan (R 2 ) diberikan untuk setiap model. Hasil yang signifikan pada α = 5% disorot dalam huruf tebal.
| Waktu | Air | Padang rumput | Sawah | Bidang | Pemandangan kota | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Karakteristik komunitas | pendidikan | F/x 2 | P | pendidikan | F/x 2 | P | pendidikan | F/x 2 | P | pendidikan | F/x 2 | P | pendidikan | F/x 2 | P | pendidikan | F/x 2 | P | R 2 |
| Keanekaragaman taksonomi | |||||||||||||||||||
| Kekayaan spesies total | 0.8 | 3.3 | 0,010 | <0,1 | <0,1 | 0.342 | <0,1 | <0,1 | 0,785 tahun | <0,1 | <0,1 | 0.488 | 0.8 | 139.3 | 0,023 | 0.6 | 9.6 | 0.198 | 0.647 |
| Kekayaan sp. spesialis hutan | 0.9 | 12.1 | <0,001 | 1.9 | 462.3 | 0,003 | <0,1 | <0,1 | 0.453 | 1.8 | 3341.9 | <0,001 | 0.9 | 5130 | <0,001 | 0.9 | 1345.3 | <0,001 | 0.845 |
| Kekayaan sp. generalis hutan | <0,1 | <0,1 | 0.942 | 0.7 | 14.2 | 0.104 | <0,1 | <0,1 | 0.793 | 2.0 | 368.0 | <0,001 | <0,1 | <0,1 | 0.302 | <0,1 | <0,1 | 0.753 | 0,567 tahun |
| Kekayaan sp. spesialis lahan terbuka | <0,1 | <0,1 | 0,935 | 1.7 | 3486.1 | <0,001 | 0.8 | 203.6 | 0,024 | 1.8 | Tahun 1868.1 | <0,001 | 1.8 | 444.2 | 0,002 | 0.9 | 1179.1 | <0,001 | 0,766 tahun |
| Spesies lainnya | 1.4 | 4.6 | 0,196 | 1.0 | 1217.4 | <0,001 | 0.8 | 18.7 | 0.114 | 0.9 | 106.0 | 0,009 | <0,1 | <0,1 | 0.691 | <0,1 | <0,1 | 0,788 tahun | 0,395 |
| Keanekaragaman fungsional | |||||||||||||||||||
| Bahasa Inggris | 0.1 | <0,1 | 0.331 | 1.3 | 316.3 | <0,001 | <0,1 | <0,1 | 0.756 | 1.6 | 79.5 | 0,001 | 0.1 | 0.1 | 0.450 | 0.8 | 27.2 | 0,033 | 0.456 |
| Februari | 1.1 | 1.0 | 0,051 tahun | 1.0 | 7.5 | 0,038 | 1.5 | 20.5 | 0,010 | <0,1 | <0,1 | 0.714 | 0.8 | 2.8 | 0.118 | 0.9 | 3.5 | 0,145 | 0.334 |
| FDI | <0,1 | <0,1 | 0,766 tahun | 0.9 | 79.4 | 0,002 | 0.8 | 19.9 | 0,046 tahun | <0,1 | <0,1 | 0,777 tahun | <0,1 | <0,1 | 0.791 | 0.9 | 121.1 | 0,001 | 0.488 |
| Bahasa Indonesia: RaoQ | <0,1 | <0,1 | 0.850 | 0.9 | 113.3 | 0,002 | 0.7 | 15.4 | 0,068 tahun | <0,1 | <0,1 | 0,985 | <0,1 | <0,1 | 0.604 | 0.9 | 150.4 | 0,001 | 0.492 |
| Keanekaragaman filogenetik | |||||||||||||||||||
| PD | 0.9 | 7.1 | <0,001 | 1.4 | 137.9 | 0,005 | <0,1 | <0,1 | 0.318 | 0.8 | 25.8 | 0,064 tahun | 0.7 | 32.4 | 0,059 tahun | 1.1 | 13.5 | 0.228 | 0.553 |
| PSV (Pekerjaan Sosial Berkelanjutan) | 1.1 | 4.8 | 0,001 | 1.5 | 206.4 | <0,001 | 0.7 | 8.9 | 0.129 | 1.1 | 98.0 | 0,001 | 0.5 | 3.9 | 0.173 | <0,1 | <0,1 | 0.488 | 0,547 tahun |
| Stabilitas temporal | |||||||||||||||||||
| Variasi seimbang | – | – | – | <0,1 | <0,1 | 0.743 | 1.4 | 5.1 | 0,021 | 0.6 | 1.3 | 0.128 | <0,1 | <0,1 | 0.413 | <0,1 | <0,1 | 0,958 | 0,075 hari |
| Gradien kelimpahan | – | – | – | 1.8 | 6.1 | 0,028 | <0,1 | <0,1 | 0.850 | <0,1 | <0,1 | 0.474 | <0,1 | <0,1 | 0.763 | 1.7 | 7.4 | 0,012 | 0,064 tahun |
Mengenai stabilitas temporal komposisi komunitas, komunitas burung yang dipelajari berkisar dari sangat stabil (komponen variasi seimbang = 0,17, komponen gradien kelimpahan = 0,01) hingga tidak stabil (komponen variasi seimbang = 0,55, komponen gradien kelimpahan = 0,21). Stabilitas temporal komunitas burung secara signifikan didorong oleh proporsi padang rumput, badan air, dan lanskap perkotaan (Tabel 1 ). Situs dengan proporsi padang rumput yang rendah menunjukkan komunitas yang lebih stabil, sedangkan situs dengan tutupan padang rumput yang lebih tinggi menunjukkan penggantian spesies yang lebih tinggi dari waktu ke waktu (Gbr. 4 ). Ketidakstabilan komposisi komunitas, didorong oleh perubahan temporal dalam kelimpahan spesies, lebih tinggi di situs dengan proporsi lanskap perkotaan yang sangat rendah atau sangat tinggi dan di situs dengan tutupan badan air menengah.

Diskusi
Dalam studi ini, kami mengeksplorasi perubahan temporal dalam keanekaragaman taksonomi, fungsional dan filogenetik komunitas burung yang menghuni lanskap Satoyama di Jepang. Satoyama merupakan lanskap yang sangat penting dalam meningkatkan keanekaragaman spesies, dan karena alasan ini, diusulkan sebagai alat untuk mempertahankan keanekaragaman hayati (Kadoya dan Washitani 2011 ). Secara khusus, kami meneliti hubungan antara perubahan keanekaragaman burung dan komposisi penggunaan lahan yang menjadi ciri Satoyama. Pertama, kami menemukan pola temporal yang signifikan secara statistik dalam metrik keanekaragaman burung yang berbeda. Secara keseluruhan, jumlah spesies burung yang menghuni Satoyama menunjukkan sedikit peningkatan dalam sepuluh tahun terakhir, tren yang juga disorot dalam kekayaan spesies spesialis hutan (Informasi pendukung). Sekilas, temuan ini dapat dilihat sebagai sesuatu yang kontras dengan penurunan keseluruhan kekayaan spesies dan tren negatif yang ditunjukkan untuk burung lahan pertanian di berbagai negara di Eropa (Chamberlain dan Fuller 2000 , Wretenberg et al. 2010 , Díaz et al. 2022 , Floigl et al. 2022 ). Namun, peningkatan sedang dalam kekayaan spesies burung spesialis hutan, yang dapat dikaitkan dengan suksesi vegetasi di lanskap Satoyama (Katayama et al. 2024 ), dapat menjadi alasan untuk perbedaan ini.
Tingkat kemerataan fungsional adalah satu-satunya ukuran keanekaragaman fungsional komunitas burung yang meningkat secara signifikan selama bertahun-tahun (Gbr. 1 ). FEve, menurut definisi, tidak berkorelasi secara matematis dengan jumlah total spesies (Mason et al. 2005 ). Kemudian, peningkatan kemerataan dalam himpunan dapat menunjukkan redundansi ekologi yang lebih tinggi, yang dapat diterjemahkan menjadi kapasitas yang lebih tinggi untuk mengatasi stresor lingkungan, untuk melawan invasi dan perubahan iklim (Villéger et al. 2008 , Haegeman et al. 2016 ). Redundansi fungsional dalam suatu himpunan meningkat ketika beberapa spesies memiliki fungsi atau sifat yang serupa. Spesies tersebut diharapkan memiliki peran yang sama dalam proses ekosistem utama (Bello et al. 2007 , Laureto et al. 2015 ). Karena alasan ini, kehilangan salah satu spesies ini seharusnya memiliki efek yang lebih kecil pada fungsi keseluruhan ekosistem (Loreau 2004 ). Meskipun demikian, peningkatan keseluruhan dalam jumlah spesies dan potensi ketahanan dalam komunitas burung lahan pertanian yang terdeteksi dalam penelitian kami terlalu kecil untuk mewakili keuntungan serius dalam hal konservasi yang efektif.
Selain itu, kami menemukan tren positif dalam keanekaragaman filogenetik komunitas burung yang menghuni lanskap Satoyama selama bertahun-tahun. Namun, peningkatan keanekaragaman filogenetik ini dapat dikaitkan dengan korelasi positif yang kuat antara PD dan jumlah spesies (Informasi pendukung). Yang lebih menarik adalah pola yang ditunjukkan oleh keterkaitan filogenetik (ukuran yang dapat dikaitkan dengan stabilitas ekosistem) (Cadotte et al. 2012 ). Kami menemukan sedikit peningkatan tetapi konstan dan signifikan dalam nilai PSV selama bertahun-tahun. Jika mengikuti hipotesis keterkaitan persaingan Darwin, kita dapat berharap bahwa dalam komunitas dengan spesies yang lebih dekat hubungannya secara filogenetik, tingkat persaingan harus lebih tinggi, bahkan jika hipotesis tersebut tidak sepenuhnya ditunjukkan atau bahkan kontras dalam beberapa komunitas burung (Sebastián-González dan Green 2017 ). Sebaliknya, spesies yang kurang terkait secara filogenetik dapat mewakili rentang ruang ekologis (atau fungsional) yang lebih luas. Dari sudut pandang ini, temuan kami yang menunjukkan peningkatan simultan dalam kekayaan spesies dan keterkaitan filogenetik dalam komunitas burung Satoyama dapat menunjukkan proses homogenisasi biotik, suatu proses yang dikenal baik untuk burung di semua lingkungan antropogenik (McKinney 2002 , Devictor et al. 2007 ).
Ketika meneliti dampak komposisi lahan yang berbeda terhadap tren keanekaragaman burung, kami menemukan bahwa setiap dimensi keanekaragaman burung merespons secara berbeda terhadap jenis penggunaan lahan yang berbeda. Secara keseluruhan, jumlah badan air merupakan penggunaan lahan utama yang memengaruhi keanekaragaman fungsional dan filogenetik di lanskap Satoyama. Cakupan badan air, yang menyediakan ceruk perairan, meningkatkan kekayaan fungsional secara keseluruhan, serta penyebaran sifat fungsional dalam komunitas burung (mungkin meningkatkan keluasan sifat berkat keberadaan beberapa spesies yang terkait dengan permukaan air). Kolam air dapat menjadi habitat yang menarik juga bagi spesies yang tidak berkembang biak. Dalam penelitian kami, 99,24% pengamatan diklasifikasikan sebagai spesies berkembang biak tertentu. Beberapa pengamatan (kurang dari 0,76% dari total) mencatat burung yang tidak diklasifikasikan sebagai spesies berkembang biak tertentu (Informasi pendukung). Sebagian besar spesies tersebut adalah bebek yang melewati musim dingin yang mungkin masih ada hingga April. Selain itu, di beberapa lokasi utara yang dingin seperti Hokkaido, beberapa bebek dapat berkembang biak. Akhirnya, beberapa burung plover dapat dicatat mengunjungi Jepang pada bulan April dan Mei sebagai migran musim panas (Fujioka et al. 2010 ). Jelas, tingkat redundansi ekologis (diukur sebagai kemerataan) menurun dengan meningkatnya cakupan badan air, karena kumpulan dengan spesies baru yang terkait dengan lingkungan tersebut tentu lebih heterogen secara fungsional. Dua ukuran keragaman filogenetik, di sisi lain, menunjukkan puncak yang sesuai dengan tingkat cakupan badan air yang sedang (misalnya bentuk U terbalik). Hasil ini menunjukkan bahwa pengelolaan pertanian yang mempertahankan petak-petak kecil dengan badan air, seperti kolam pertanian dan parit, dapat membantu konservasi keanekaragaman hayati dengan meningkatkan warisan evolusi yang diwakili oleh kumpulan spesies (Deguchi et al. 2020 ).
Secara keseluruhan, kekayaan spesies hanya berasosiasi secara signifikan dan negatif dengan proporsi ladang, dengan nilai minimum tercapai ketika tutupan ladang lebih tinggi dari 60% lanskap Satoyama. Untuk beberapa ukuran keanekaragaman hayati, proporsi sawah ditemukan memiliki efek yang signifikan (misalnya bentuk-u terbalik untuk generalis hutan, negatif untuk spesies spesialis hutan, positif untuk spesialis lahan terbuka dan sedikit positif untuk spesies lain), sementara proporsi lahan kering lainnya memiliki efek negatif. Ini menunjukkan bahwa keanekaragaman burung dapat meningkat atau menurun tergantung pada bagaimana lahan pertanian dikelola. Sawah diketahui mendukung banyak spesies burung air seperti plover, bebek, angsa, kuntul dan ibis secara global (Pierluissi 2010 , Deguchi et al. 2020 ). Di sisi lain, lahan kering seperti gandum dan sayuran tampaknya mendukung lebih sedikit spesies, seperti burung skylark dan burung pegar. Kekayaan spesialis hutan menurun secara signifikan dengan tutupan perkotaan, sementara kekayaan spesialis lahan terbuka menurun dengan jenis tutupan penggunaan lahan tersebut. Selain itu, tutupan air memengaruhi kekayaan masing-masing jenis spesies secara berbeda, meningkatkan jumlah spesies spesialis lahan terbuka dan spesies lainnya sekaligus mengurangi kekayaan spesies spesialis hutan secara keseluruhan.
Keseragaman dan penyebaran fungsional cenderung lebih rendah ketika meningkatkan tutupan padang rumput, sementara kekayaan fungsional, penyebaran, dan entropi kuadrat Rao meningkat secara signifikan ketika meningkatkan cakupan area perkotaan (meskipun selalu di bawah 40% dari total). Hasil ini dapat mencerminkan ketersediaan besar relung baru (terhuni), yang diperkenalkan oleh keberadaan area buatan manusia, yang sering kali menarik spesies yang lebih generalis atau toleran terhadap perkotaan (Callaghan et al. 2019 ). Yang penting, sebagian besar lokasi survei dalam studi ini terletak di daerah pedesaan atau pinggiran kota, sehingga kelas penggunaan lahan yang didefinisikan sebagai ‘perkotaan’ terutama mengacu pada rumah dan kebun tradisional, yang sering digunakan oleh beberapa spesies Satoyama seperti burung pipit pohon Eurasia dan burung pipit hijau Oriental.
Akhirnya, ketika memeriksa stabilitas temporal komunitas burung, kami menemukan bahwa penggantian spesies dari waktu ke waktu dalam komunitas burung meningkat ketika lanskap Satoyama dicirikan oleh tutupan padang rumput yang lebih besar (misalnya variasi yang sangat seimbang). Lebih jauh lagi, tingkat ketidakstabilan komunitas burung yang lebih tinggi (misalnya gradien kelimpahan atau kehilangan bersih beberapa individu dari waktu ke waktu) terutama diperkirakan dalam kumpulan spesies yang menghuni daerah dengan tutupan air sedang atau tutupan daerah perkotaan yang sangat rendah atau sangat tinggi. Efek potensial dari perubahan iklim bukanlah fokus studi kami dan dapat menjadi keterbatasan, yang menunjukkan tantangan di masa depan. Kami tahu bahwa variabilitas iklim dapat mengubah distribusi spesies dan kumpulan spesies yang dihasilkan (Hillebrand dan Blenckner 2002 , Princé et al. 2013 ). Di daerah pegunungan Eropa, misalnya, disarankan bahwa pemanasan global yang dikombinasikan dengan tekanan antropogenik dapat berdampak negatif pada kelimpahan beberapa spesies burung (Lehikoinen et al. 2019 ). Di Jepang, dampak perubahan iklim telah dikaitkan dengan potensi kontraksi dalam rentang distribusi beberapa spesies raptor (Condro et al. 2022 ), dampak yang dapat serupa untuk banyak spesies burung lainnya (Huntley et al. 2007 ). Pengurangan dalam rentang distribusi burung karena perubahan iklim dapat menyebabkan pemiskinan fungsi dan layanan ekosistem (Stewart et al. 2022 ). Selain itu, dampak perubahan iklim dapat sangat terasa bagi burung yang bermigrasi, yang memengaruhi fenologi kejadian musiman (Kobori et al. 2012 , Seri dan Rahman 2021 ). Kami berharap bahwa pemanasan di Jepang dapat menghasilkan dampak yang lebih kuat pada pergantian temporal komunitas burung yang menghuni area terbuka seperti padang rumput atau lokasi yang sangat perkotaan daripada di area hutan (Lauck et al. 2023 , Katayama et al. 2024 ).
Meskipun kami menemukan tren yang sedikit meningkat dalam keanekaragaman burung, heterogenitas habitat lanskap Satoyama telah hilang karena depopulasi manusia, dan pengabaian lahan terkait dapat dikaitkan dengan pengurangan banyak spesies kupu-kupu di Jepang (Sugimoto et al. 2022 ). Temuan kami menunjukkan bahwa jika depopulasi lebih lanjut, pengabaian dan hilangnya habitat terbuka seperti sawah dan badan air terjadi di masa depan, keanekaragaman burung dapat menurun. Keluaran penelitian utama kami menyoroti pentingnya lanskap Satoyama yang mendukung konservasi keanekaragaman hayati. Namun, komposisi relatif jenis penggunaan lahan memainkan peran penting dalam membentuk berbagai komponen keanekaragaman taksonomi, fungsional dan filogenetik dalam kumpulan burung, serta stabilitas temporal mereka. Kami mengklaim bahwa strategi perencanaan konservasi yang lebih akurat harus memperhatikan setiap dimensi keanekaragaman burung, terutama pada tren temporal yang mencirikan kumpulan burung yang menghuni lanskap kuno dan tradisional Jepang.