
ABSTRAK
Telomer, tutup pelindung di ujung kromosom linear, sering ditemukan memendek seiring bertambahnya usia. Panjang telomer umumnya diukur pada populasi liar untuk menyelidiki perubahan terkait usia dalam integritas somatik dan dianggap sebagai ciri khas penuaan. Meskipun menarik, tidak ada gambaran yang jelas mengenai perbedaan jenis kelamin dalam panjang telomer atau tingkat pengurangan antar spesies. Kelelawar sangat menarik dalam studi penuaan dan telomer, karena umur panjangnya yang luar biasa dan tidak adanya pengurangan telomer terkait usia yang diamati pada beberapa spesies. Selain itu, banyak spesies kelelawar menunjukkan bukti perbedaan jenis kelamin dalam umur panjang. Namun, sedikit penelitian telomer kelelawar yang mencakup kedua jenis kelamin. Kami mengumpulkan DNA dari jantan dan betina yang ditangkap di alam liar dari kelelawar hidung tombak besar yang sangat poligini, Phyllostomus hastatus , di mana mortalitas sangat bias jantan, dan mengukur panjang telomer relatif. Kami menemukan bahwa, meskipun telomer lebih pendek pada kelelawar yang lebih tua, tidak ada bukti telomer yang lebih pendek pada kelelawar jantan. Faktanya, kelelawar jantan cenderung memiliki telomer yang lebih panjang. Hal ini bertentangan dengan prediksi kami tentang telomer yang lebih pendek pada jenis kelamin yang berumur pendek, tetapi tidak sepenuhnya tidak terduga mengingat pengamatan lain, termasuk pengamatan telomer yang lebih pendek pada spesies yang berumur lebih panjang.
1 Pendahuluan
Bahasa Indonesia : Berbagai taksa hewan menunjukkan perbedaan jenis kelamin dalam rentang hidup, dengan individu dari satu jenis kelamin secara konsisten hidup lebih lama dari yang lain (Lemaître et al. 2020 ; Marais et al. 2018 ). Sementara perbedaan-perbedaan ini pada akhirnya mungkin timbul dari strategi reproduksi yang berbeda dari jantan dan betina (Tidière et al. 2015 ), penyebab langsung dari perbedaan jenis kelamin dalam umur panjang kurang dipahami (Austad dan Fischer 2016 ) dan bervariasi antar spesies, yang melibatkan kombinasi faktor intrinsik dan ekstrinsik (Bronikowski et al. 2022 ). Faktor intrinsik yang menarik secara luas dalam konteks penuaan adalah panjang telomer. Telomer adalah wilayah DNA berulang dan non-coding yang menutupi ujung kromosom linear, melindunginya terhadap kesalahan replikasi DNA dan kerusakan oksidatif, tetapi memendek selama pembelahan sel (Aubert dan Lansdorp 2008 ). Daerah-daerah ini dengan demikian sering, meskipun tidak selalu, memendek seiring bertambahnya usia; pengurangan telomer dianggap sebagai satu dari lima ciri utama penuaan (López-Otín et al. 2023 ), atau, sebagai alternatif, biomarker integritas atau redundansi somatik (Boonekamp et al. 2013 ; Wood dan Young 2019 ). Pemendekan telomer secara progresif dianggap mewakili pertahanan terhadap replikasi sel yang tidak terkendali yang terkait dengan pertumbuhan tumor (Maciejowski dan de Lange 2017 ; Schmutz et al. 2020 ), tetapi telomer yang lebih pendek juga dikaitkan dengan berbagai penyakit dan gangguan serta peningkatan risiko kematian (Savage dan Bertuch 2010 ; Wilbourn et al. 2018 ), menunjukkan adanya trade-off yang mungkin dipengaruhi oleh strategi riwayat hidup.
Di antara vertebrata, ada tren umum bagi telomer untuk memendek seiring bertambahnya usia, atau bagi individu yang lebih tua untuk memiliki telomer yang lebih pendek. Namun, pola ini bervariasi antara spesies dan memiliki pengecualian penting (Remot et al. 2022 ), termasuk spesies yang menunjukkan bukti pemanjangan telomer seiring bertambahnya usia (Tissier et al. 2022 ). Dalam spesies, jenis kelamin merupakan salah satu kemungkinan sumber heterogenitas dalam pola pengurangan telomer, dan perbedaan jenis kelamin dalam panjang telomer di seluruh taksa hewan telah menjadi sumber perdebatan. Populasi manusia di seluruh dunia menunjukkan mortalitas yang bias terhadap laki-laki (Austad dan Fischer 2016 ), dan laki-laki biasanya memiliki telomer yang lebih pendek daripada wanita (Gardner et al. 2014 ). Pola yang konsisten telah diperhatikan dalam taksa vertebrata lain, terutama mamalia, dengan penulis menyarankan bahwa jantan cenderung menunjukkan telomer yang lebih pendek atau tingkat pengurangan telomer yang lebih besar terkait usia (Barrett dan Richardson 2011 ). Namun, sebuah meta-analisis baru-baru ini menantang pandangan bahwa jantan cenderung memiliki telomer yang lebih pendek, tidak menemukan bukti perbedaan jenis kelamin yang konsisten dalam panjang telomer di seluruh mamalia, atau vertebrata secara umum, atau hubungan apa pun antara perbedaan jenis kelamin dalam panjang atau umur telomer (Remot et al. 2020 ). Analisis ini tidak menguji perbedaan jenis kelamin dalam tingkat pengurangan telomer, yang dapat menjadi indikasi tingkat penuaan yang berbeda, dan mungkin merupakan biomarker yang lebih informatif (Wood dan Young 2019 ), tetapi datanya relatif jarang. Seperti yang dicatat oleh para penulis, jika jenis kelamin memiliki panjang telomer yang sama di awal masa dewasa, tetapi telomer mereka memendek pada tingkat yang berbeda, maka perbedaan jenis kelamin akan paling menonjol pada individu yang lebih tua, yang kurang terwakili dalam banyak penelitian. Selain itu, jika usia tidak diketahui, dan usia memengaruhi panjang telomer, perbandingan dapat membingungkan karena perbedaan jenis kelamin dalam distribusi usia.
Kelelawar sangat menarik dalam studi penuaan dan panjang telomer (Power et al. 2022 ; Teeling 2021 ). Hal ini sebagian disebabkan oleh umur panjang mereka yang mengesankan dibandingkan dengan mamalia berukuran sama, dengan kelelawar memiliki umur rata-rata yang disesuaikan dengan massa tubuh sekitar 3,5 kali lipat dari mamalia plasenta berukuran sama lainnya (Austad 2010 ; Wilkinson dan Adams 2019 ). Studi sebelumnya telah menunjukkan pemeliharaan telomer mungkin berperan dalam perpanjangan umur beberapa spesies kelelawar (Foley et al. 2018 ). Kelelawar juga sering menunjukkan bukti perbedaan jenis kelamin dalam umur, dengan jantan atau betina dari spesies yang berbeda hidup lebih lama dari jenis kelamin lainnya, terkadang secara dramatis (Adams et al. 2025 ; Austad 2010 ). Namun, penelitian tentang panjang telomer pada kelelawar jarang menyertakan jumlah jantan dan betina yang sama dari seluruh rentang usia (Foley et al. 2018 ; Ineson et al. 2020 ; Power et al. 2022 ) (tetapi lihat (Power et al. 2023 )), mungkin karena sulitnya menangkap kedua jenis kelamin dalam jumlah yang cukup untuk banyak spesies.
Dalam studi saat ini, kami menguji apakah panjang telomer dikaitkan dengan jenis kelamin atau usia pada populasi kelelawar berhidung tombak besar Phyllostomus hastatus di Trinidad , spesies yang sangat poligami di mana jantan memiliki kematian yang jauh lebih awal. Jantan dari spesies ini bersaing untuk dan mempertahankan kelompok harem betina (biasanya dengan 13–23 betina per harem) (McCracken dan Bradbury 1981 ). Mungkin sebagai hasil dari riwayat hidup yang berbeda yang terkait dengan sistem perkawinan harem-poligami ini, jantan hidup sekitar setengah lebih lama dari betina (Adams et al. 2025 ), yang telah dilaporkan hidup hingga 22 tahun (Wilkinson dan Adams 2019 ). Kami mengekstrak DNA dari biopsi sayap kelelawar jantan dan betina yang ditangkap di alam liar, yang kami gunakan untuk mengukur panjang telomer relatif dengan qPCR dan yang juga digunakan dalam studi paralel untuk memperkirakan usia kronologis dari data metilasi DNA menggunakan jam metilasi yang dipublikasikan (Wilkinson et al. 2021 ). Kami memperkirakan bahwa panjang telomer akan berasosiasi negatif dengan usia yang diestimasikan dan bahwa asosiasi negatif ini akan lebih kuat pada jantan, karena mortalitas mereka yang lebih awal yang menunjukkan potensi percepatan kemunduran somatik. Dengan memanfaatkan spesies dengan perbedaan jenis kelamin yang jelas dalam umur panjang, studi kami merupakan kontribusi penting untuk menguji prediksi panjang telomer yang berkurang dan pola pemendekan telomer yang lebih besar terkait usia pada jenis kelamin yang berumur lebih pendek, yang buktinya saat ini masih samar-samar.
2 Metode
2.1 Pengambilan sampel
Sampel jaringan dikumpulkan pada bulan Januari (menjelang akhir musim kawin) tahun 2023 dan 2024. Kelelawar ditangkap dari tiga lokasi di Trinidad: formasi gua alami di Gunung Tamana (10.4711° LU, 61.1958° BB; N = 111 dari tahun 2023 dan 2024) dan lembah Caura (10.7019° LU, 61.3614° BB; N = 28 dari tahun 2023), dan bangunan penyimpanan dingin yang terbengkalai di Cumuto (10.5983° LU, 61.2117° BB; N = 22 dari tahun 2024). Pada siang hari, P. hastatus bertengger dalam kelompok yang terdiri dari beberapa betina (dengan atau tanpa anak) yang dijaga oleh satu jantan harem, atau beberapa jantan ‘bujangan’ atau bawahan. Untuk menangkap kelompok, yang menghuni cekungan kecil di langit-langit gua, kami menggunakan perangkap ember, yang bagian bawahnya diganti dengan keranjang cucian berbahan nilon untuk mencegah kelelawar melarikan diri. Perangkap ember ini dinaikkan untuk melingkupi kelompok, menyebabkan kelelawar terbang atau jatuh ke dalam keranjang. Kelelawar ditempatkan satu per satu di dalam kantong hingga diproses, di mana mereka ditimbang hingga 0,1 g terdekat, lengan bawah kiri mereka diukur hingga 0,01 mm terdekat menggunakan jangka sorong digital, dan keausan gigi mereka (korelasi usia) dinilai pada skala 1 (tidak aus) hingga 5 (sangat aus) dengan peningkatan 0,5 (McCracken dan Bradbury 1981 ). Akhirnya, kami mengambil biopsi sayap sepanjang 4 mm dari membran setiap sayap kelelawar dewasa (Power et al. 2021 ). Biopsi membran sayap bersifat minimal invasif, dan cepat sembuh (Faure et al. 2009 ). Jaringan disimpan dalam pelindung Zymo DNA/RNA dan dibekukan pada suhu -20°C, hingga ekstraksi DNA menggunakan kit Zymo Quick-DNA Miniprep Plus. Semua kelelawar dilepaskan kembali ke tempat bertenggernya. DNA yang dimurnikan disimpan dalam Zymo DNA Elution Buffer pada suhu -20°C. Konsentrasi DNA diukur menggunakan uji fluoresensi Qubit sensitivitas tinggi dan selanjutnya diencerkan menjadi 2 ng/μl untuk digunakan dalam reaksi qPCR. Karena jumlah DNA yang tersisa rendah, kami tidak dapat menilai kemurnian dan integritas DNA secara sistematis. Dalam sampel DNA yang diekstraksi dalam kondisi yang sama, kami mengamati rasio 260/280 ~1,8 pada Nanodrop ND-1000 dengan andal, dan karena semua sampel dikumpulkan dan diekstraksi menggunakan protokol yang sama, setiap variasi kemurnian harus didistribusikan secara acak di seluruh sampel. Kami menguji integritas DNA menggunakan elektroforesis gel untuk subset 26 sampel dan mengamati pita bening > 10Kb dengan noda minimal.
2.2 Uji Panjang Telomer
Kami memperkirakan panjang telomer relatif (rTL) untuk setiap sampel menggunakan qPCR, seperti dalam penelitian rTL sebelumnya menggunakan DNA yang diekstraksi dari biopsi sayap pada spesies kelelawar (Foley et al. 2018 ; Ineson et al. 2020 ; Power et al. 2022 , 2023 ). Kami menggunakan sekuens primer Tel1b dan Tel2b (Cawthon 2002 ) untuk mengamplifikasi sekuens telomerik, dan primer berdasarkan genom P. hastatus (Santillán et al. 2021 ) untuk mengamplifikasi faktor neurotropik yang berasal dari otak BDNF (Tabel S1 ), yang kami gunakan sebagai gen referensi salinan tunggal (lihat (Foley et al. 2018 )). Kami melakukan pengujian qPCR menggunakan PowerUp SYBR green Master Mix untuk qPCR (Applied Biosystems) pada Roche 480 Lightcycler. Rincian volume reaksi qPCR dan pengaturan thermocycler ada di Tabel S2 . Kami menggunakan seri pengenceran untuk menguji efisiensi amplifikasi, yang diperkirakan sebesar 103,66% (95% CI: 96,64%, 111,75%) dan 95,63% (95% CI: 87,95%, 104,76%) untuk primer telomer dan BDNF , masing-masing (Gambar S1 ).
Kami menjalankan setiap kombinasi sampel dan primer dalam rangkap tiga, dengan kedua primer disertakan pada plat yang sama untuk setiap sampel. Semua plat menyertakan sampel kalibrator dan kontrol tanpa templat (juga dijalankan dalam rangkap tiga). Kami menghitung rTL mengikuti Pfaffl (Pfaffl 2001 ). Kami menjalankan setiap konfigurasi plat dua kali untuk memperhitungkan variasi antar-plat dan menggunakan pengukuran rTL rata-rata dalam analisis kami. Kami menjalankan subset sampel ( N = 15) pada tiga plat karena dua estimasi rTL pertama tidak sama. Kami menilai rentang nilai C p untuk setiap rangkap tiga. Jika rentangnya lebih besar dari 1 dan ada outlier yang jelas, kami menghapus outlier tersebut. Jika tidak, kami mempertahankan nilai rangkap tiga dengan asumsi bahwa rata-rata adalah estimasi yang wajar tetapi memeriksa bahwa interpretasi hasil tidak terpengaruh oleh penghapusan rangkap tiga dengan rentang nilai > 1. Kami melakukan prosedur yang sama untuk pengukuran rTL sampel yang dijalankan pada tiga plat. Sampel yang estimasi rTL replikasinya memiliki koefisien variasi lebih besar dari 50% ( N = 7) dikecualikan dari analisis, karena visualisasi menunjukkan bahwa sampel tersebut memiliki nilai rTL rata-rata dengan keyakinan rendah. Kami menjalankan pengujian dalam dua kelompok, dengan 86 sampel pertama dikumpulkan pada Januari 2023 dan diuji pada Juli/Agustus 2023, dan 57 sampel kedua dikumpulkan pada Januari 2024 dan diuji pada Mei/Juni 2024. Sampel referensi yang sama (ID Band 2311) digunakan di kedua kelompok.
Kemampuan mengulang nilai C p primer yang ditransformasikan log 2 untuk triplikat dan ukuran rTL dari plat duplikat dihitung menggunakan rptR (Stoffel et al. 2017 ), dengan asumsi distribusi kesalahan Gaussian dan dengan 1000 bootstrap. Kemampuan mengulang intraplat dari nilai titik persilangan yang ditransformasikan log2 untuk ukuran telomer dan BDNF juga dihitung menggunakan rptR, dengan kovariat ID plat. Kemampuan mengulang intraplat dari nilai titik persilangan untuk sekuens telomer dan BDNF masing-masing adalah 0,933 (95% CI = 0,912, 0,951) dan 0,945 (0,925, 0,958), sedangkan kemampuan mengulang interplat dari ukuran rTL adalah 0,767 (0,685, 0,827) (Gambar S3 ).
2.3 Estimasi Usia
Data metilasi dihasilkan sebagai bagian dari studi paralel (Adams et al. 2025 ) dengan mengirimkan DNA yang dimurnikan ke Epigenetic Clock Development Foundation (Torrance, CA), tempat sampel mengalami konversi bisulfit dan dihibridisasi ke array HorvathMammal40. Usia individu diperkirakan menggunakan jam “semua kelelawar” yang diterbitkan, yang dirancang untuk memperkirakan usia kronologis secara akurat untuk setiap spesies kelelawar berdasarkan profil metilasi pada jaringan kulit (Wilkinson et al. 2021 ). Kami mencatat bahwa sampel dari betina yang tampak lebih tua (berdasarkan keausan gigi atau catatan pita) terkadang diprioritaskan untuk profil metilasi dalam studi paralel, yang mengakibatkan kurangnya representasi betina muda dalam sampel kami (Adams et al. 2025 ). Ini tidak berlaku untuk jantan, yang hampir semua sampelnya disertakan dalam uji metilasi. Dalam pengujian perbedaan jenis kelamin dalam panjang telomer, kami memperhitungkan bias pengambilan sampel ini dengan menggunakan prosedur subsampling untuk memilih pria dan wanita yang berusia sama, yang dijelaskan secara rinci di bawah ini.
Di antara 143 kelelawar yang disertakan dalam analisis akhir, 138 di antaranya diperkirakan usianya berdasarkan profil metilasi. Untuk sisanya, kami mengetahui usia dua kelelawar berdasarkan penangkapan saat masih muda, dan kami memperkirakan usia empat kelelawar betina lainnya berdasarkan regresi khusus betina dari skor keausan gigi pada perkiraan usia (Perkiraan usia = 3,0664 × keausan gigi—0,4828; R 2 = 0,693, F 1,74 = 166,9, p < 0,001) (Wilkinson et al. 2024 ).
2.4 Analisis Statistik
Data dianalisis menggunakan R (v4.3.1) (Tim 2020 ). Dimorfisme ukuran seksual di antara ukuran panjang lengan bawah dan berat badan (transformasi-z) diuji menggunakan regresi linier dengan prediktor jenis kelamin dan populasi. Untuk berat badan, kami menyertakan efek kuadrat dari perkiraan usia (berpusat pada rata-rata) menggunakan fungsi poli dalam R, dan interaksinya dengan jenis kelamin, karena plot eksplorasi menunjukkan hubungan non-linier yang berbeda antara jenis kelamin (Gambar S2 ).
Hubungan antara estimasi rTL rata-rata (dirata-ratakan di seluruh pengulangan lari plat) per sampel dan variabel prediktor jenis kelamin, usia, panjang lengan bawah, dan populasi diuji menggunakan regresi linier, dengan variabel respons log 2 -ditransformasikan untuk meningkatkan kenormalan residual, kemudian ditransformasikan z (Verhulst 2020 ). Kami juga memasukkan sebagai prediktor: tahun pengambilan sampel (2023 atau 2024); interval antara pengumpulan sampel dan uji qPCR (Power et al. 2023 ); dan interval antara tanggal pengambilan sampel kalibrator dan uji qPCR. Ketiga prediktor ini kolinear tetapi dimasukkan untuk memperhitungkan variasi pengganggu tambahan antara pengukuran rTL (Morrissey dan Ruxton 2018 ). Variabel kontinu berpusat pada rata-rata. Kami awalnya memasukkan suku kuadrat untuk variabel kontinu, tetapi menghapusnya karena suku orde kedua tidak mendekati signifikansi pada p < 0,05. Sebelas individu diambil sampelnya di kedua tahun tersebut. Upaya untuk memasukkan ID sebagai intersepsi acak guna memperhitungkan pengukuran berulang menyebabkan kecocokan model tunggal, jadi kami mengecualikan sampel Januari 2023 dari individu-individu ini dalam analisis. Mengambil pendekatan alternatif dengan menghapus sampel Januari 2024 dari analisis kami tidak berpengaruh pada interpretasi hasil.
Kami menyelidiki kerentanan hasil kami terhadap ketidakpastian dalam estimasi rTL atau usia menggunakan analisis bootstrap. Kami mengambil estimasi median absolute error (MAE) untuk estimasi usia sebesar 0,449 dari (Adams et al. 2025 ) dan menghitung MAE dari estimasi rTL berulang kami sebesar 0,243. Dalam setiap 10.000 bootstrap, kami menambahkan nilai acak ke estimasi usia dan rTL dengan distribusi seragam yang dibatasi antara obs-AE dan obs + MAE, lalu menjalankan kembali analisis kami.
Model dengan dan tanpa istilah interaksi diuji menggunakan jumlah kuadrat tipe III dan II, masing-masing. Kami menggunakan paket R ggeffects untuk memplot, dari model panjang rTL, nilai prediksi yang disesuaikan di seluruh jenis kelamin dan usia, dengan mengendalikan istilah non-fokus menggunakan marginalisasi ‘empiris’ (Lüdecke 2018 ). Data tersedia sebagai Informasi Pendukung— S1 .
3 Hasil
3.1 Ukuran Tubuh
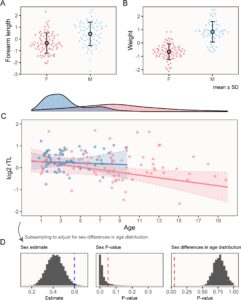
Kami mengamati dimorfisme ukuran seksual yang substansial, konsisten dengan temuan sebelumnya (Adams et al. 2020 ), dan dengan prediksi untuk sistem perkawinan yang sangat poligami (Clutton-Brock 1985 ). Laki-laki memiliki ukuran tubuh struktural yang lebih besar (Gambar 1A ) dan jauh lebih berat (Gambar 1B ) di semua usia. Berat badan juga dikaitkan dengan istilah kuadrat dari perkiraan usia, yang berbeda antara jenis kelamin (Tabel 1 , Gambar S2 ). Distribusi perkiraan usia berbeda secara substansial antara jenis kelamin (Gambar 1C ), dengan usia maksimum perempuan 19,80 tahun (median = 8,30, IQR [5,70, 11,25]), versus 9,70 tahun untuk laki-laki (median = 3,00, IQR [1,80, 4,20]) (Gambar 1C ). Sampel tidak acak berkenaan dengan distribusi usia perempuan (lihat Metode), jadi kami tidak membandingkan secara statistik distribusi usia yang diestimasikan dalam sampel kami. Meskipun demikian, perbedaan jenis kelamin dalam usia maksimum konsisten dengan pengamatan umur panjang yang cenderung condong pada perempuan dalam spesies ini (Adams et al. 2025 ; Wilkinson et al. 2024 ) dan kami mengambil sampel dari rentang penuh usia dewasa pada kedua jenis kelamin.
| Tanggapan | Prediktor | Koefisien ± SE | F | df | P |
|---|---|---|---|---|---|
| Panjang lengan bawah | Mencegat | -0,500 ± 0,189 | |||
| Jenis Kelamin (L) | 0,860 ± 0,152 | 31.951 | 1 | < 0,001 | |
| Populasi | 6.806 | 2 | 0,002 | ||
| Caura | -0,290 ± 0,193 | ||||
| Hanya saja | -0,800 ± 0,220 | ||||
| Sisa | ± 0,888 | 139 | |||
| Berat badan | Mencegat | -0,831 ± 0,081 | |||
| Jenis Kelamin (L) | 1.896 ± 0.282 | 45.274 | 1 | < 0,001 | |
| Populasi | 3.654 | 2 | 0,031 | ||
| Caura | 0,328 ± 0,128 | ||||
| Hanya saja | 0,174 ± 0,143 | ||||
| Perkiraan usia | 3.554 | 2 | 0,035 | ||
| Perkiraan usia | 2.021 ± 0.843 | ||||
| Perkiraan usia 2 tahun | -1,376 ± 0,720 | ||||
| Jenis Kelamin × Estimasi Usia 2 | 6.282 | 2 | 0,002 | ||
| Jenis KelaminP:PerkiraanUsia | 2.558 ± 5.960 | ||||
| Jenis KelaminP:Perkiraan umur 2 | -4,171 ± 4,193 | ||||
| Sisa | ± 0,577 tahun | 135 |
3.2 Panjang Telomer Relatif
Seperti yang diprediksi, panjang telomer (dirata-ratakan di seluruh pelat replikasi) berasosiasi negatif dengan usia dalam model lengkap, termasuk kedua jenis kelamin, dari sampel penampang kami (Tabel 2 ). Namun, yang mengejutkan, telomer pria diperkirakan lebih panjang meskipun mortalitas mereka lebih awal. Mengingat distribusi usia yang berbeda pada pria dan wanita, kami tidak menyesuaikan interaksi jenis kelamin menurut usia dalam model lengkap tetapi menjalankan model khusus jenis kelamin yang melaporkan asosiasi negatif signifikan antara usia dan panjang telomer hanya di seluruh sampel wanita, mungkin dipengaruhi oleh ukuran sampel yang lebih besar ( N = 80, hingga 63 untuk pria) dan rentang usia yang lebih luas (karena wanita hidup hingga dua kali lebih lama dari pria) (Tabel 2 ).
| Prediktor | Koefisien ± SE | F | df | P | |
|---|---|---|---|---|---|
| Model lengkap ( N = 143) | Mencegat | 11.323 ± 5.039 | |||
| Panjang lengan bawah | -0,060 ± 0,048 | 1.568 | 1 | 0.213 | |
| Perkiraan usia | -0,054 ± 0,022 | 5.981 | 1 | 0,016 | |
| Jenis Kelamin (L) | 0,491 ± 0,205 | 5.705 | 1 | 0,018 | |
| Populasi | 0,983 | 2 | 0,377 tahun | ||
| Caura | -0,311 ± 0,265 | ||||
| Hanya saja | -0,242 ± 0,285 | ||||
| Tahun sampel (Jan_2024) a | -29.492 ± 12.588 | 5.488 | 1 | 0,021 | |
| Durasi penyimpanan sampel a | -0,098 ± 0,035 | 7.879 | 1 | 0,006 | |
| Durasi penyimpanan sampel kalibrator a | 0,081 ± 0,035 | 5.281 | 1 | 0,023 | |
| Sisa | ± 0,896 | 133 | |||
| Betina ( N = 80) | Mencegat | 12.143 ± 10.126 | |||
| Panjang lengan bawah | -0,048 ± 0,078 | 0.390 | 1 | 0,534 tahun | |
| Perkiraan usia | -0,061 ± 0,028 | 4.737 | 1 | 0,033 | |
| Populasi | 1.033 | 2 | 0.361 | ||
| Caura | -0,535 ± 0,405 | ||||
| Hanya saja | 0,215 ± 0,467 | ||||
| Tahun sampel (Jan_2024) a | -25.839 ± 21.248 | 1.479 | 1 | 0.228 | |
| Durasi penyimpanan sampel a | -0,096 ± 0,058 | 2.756 | 1 | 0.101 | |
| Durasi penyimpanan sampel kalibrator a | 0,068 ± 0,060 | 1.307 | 1 | 0.257 | |
| Sisa | ± 1.043 | 71 | |||
| Laki-laki ( N = 63) | Mencegat | 6.666 ± 4.408 | |||
| Panjang lengan bawah | -0,054 ± 0,058 | 1.304 | 1 | 0,355 | |
| Perkiraan usia | -0,016 ± 0,050 | 0,061 tahun | 1 | 0.747 | |
| Populasi | 1.584 | 2 | 0.304 | ||
| Caura | 0,021 ± 0,322 | ||||
| Hanya saja | -0,541 ± 0,348 | ||||
| Tahun sampel (Jan_2024) a | -21.015 ± 14.419 | 1.778 | 1 | 0.151 | |
| Durasi penyimpanan sampel a | -0,065 ± 0,041 | 2.197 | 1 | 0.114 | |
| Durasi penyimpanan sampel kalibrator a | 0,060 ± 0,040 | 1.862 | 1 | 0,144 tahun | |
| Sisa | ± 0,680 | 55 |
Catatan: Model lengkap memiliki R 2 sebesar 0,243 ( F 8,133 = 5,336, p < 0,001). Model yang hanya melibatkan perempuan dan laki-laki memiliki nilai R 2 sebesar 0,254 ( F 7,72 = 3,453, p = 0,003) dan 0,178 ( F 7,55 = 1,702, p = 0,073). a Istilah-istilah ini kolinear dan dimasukkan untuk memperhitungkan variasi perancu antara sampel, tetapi estimasi yang sesuai tidak boleh ditafsirkan secara individual (lihat Tabel S5–S7 ).
Masalah potensial dengan interpretasi bahwa laki-laki memiliki telomer yang lebih panjang, berdasarkan signifikansi istilah jenis kelamin dalam model lengkap, adalah bahwa ada perbedaan yang jelas dalam distribusi usia antara laki-laki dan perempuan dalam sampel kami (Gambar 1C ) yang dapat mengacaukan perbandingan mereka. Untuk menyelidiki apakah perbedaan jenis kelamin yang diamati dalam rTL merupakan artefak dari distribusi usia yang berbeda, kami melakukan prosedur subsampling berulang untuk menguji perbedaan jenis kelamin dalam panjang telomer relatif di seluruh subsampel acak di mana jenis kelamin memiliki distribusi usia yang sama. Singkatnya, kami mempertahankan, untuk setiap laki-laki dalam sampel kami, satu perempuan dalam usia 1 tahun (jika ada, jika tidak, laki-laki itu dibuang dari sampel). Laki-laki dilaporkan memiliki telomer yang secara signifikan ( p <0,05) lebih panjang dalam 91,99% dari 10.000 sampel acak, mendukung interpretasi awal, meskipun besarnya perbedaan berkurang dibandingkan dengan model lengkap (Gambar 1D ). Singkatnya, kami tidak mengamati dukungan yang jelas untuk prediksi percepatan pengurangan telomer atau telomer yang lebih pendek pada P. hastatus jantan yang berumur relatif pendek , dan hasil kami secara langsung bertentangan dengan prediksi terakhir.
Baik asosiasi dengan jenis kelamin maupun usia, yang dilaporkan oleh model lengkap, tampaknya tidak dipengaruhi oleh ketidakpastian dalam ukuran rTL atau usia yang diestimasikan (Gambar S5 ). Kami mengamati efek signifikan dari variasi dalam durasi penyimpanan sampel dan kalibrator, yang kolinear dan berbeda antar tahun (Tabel 2 ). Penghapusan istilah kolinear tidak memengaruhi interpretasi biologis (Tabel S3–S5 ). Karena variasi teknis ini, kami tidak dapat secara langsung membandingkan ukuran rTL untuk 11 kelelawar yang diambil sampelnya pada tahun 2023 dan 2024, tetapi perbandingan residual regresi antar tahun tidak menunjukkan arah perubahan yang jelas (yaitu, perolehan atau kehilangan rTL yang konsisten) (Gambar S4 ).
4 Diskusi
Jenis kelamin adalah sumber variasi penting dalam tingkat kematian dan penuaan pada populasi hewan liar (Bronikowski et al. 2022 ; Marais et al. 2018 ), dan pengurangan telomer dianggap oleh beberapa orang sebagai ciri penuaan atau penuaan (López-Otín et al. 2023 ). Apakah jenis kelamin berbeda dalam panjang telomer, secara konstitutif atau melalui riwayat hidup yang berbeda, tidak jelas. Seperti banyak spesies kelelawar, P. hastatus memiliki umur maksimum yang lebih panjang daripada yang diprediksi untuk mamalia seukurannya (Wilkinson dan Adams 2019 ), namun jantan hidup sekitar setengah lebih lama dari betina. Seperti yang dilaporkan sebelumnya, jantan juga lebih besar (McCracken dan Bradbury 1981 ), dengan ukuran kemungkinan merupakan faktor penting yang memengaruhi kemampuan mereka untuk bersaing dan mempertahankan kelompok besar betina, dan berpotensi juga umur mereka yang relatif pendek (Clutton-Brock et al. 1985 ). Sementara kami mengamati variasi signifikan terkait usia dalam panjang telomer di seluruh sampel kami, kami tidak mengamati telomer yang lebih panjang pada wanita dan, pada kenyataannya, pola ini tampaknya terbalik, dengan pria cenderung memiliki telomer yang lebih panjang.
Berbagai faktor telah diusulkan untuk berkontribusi pada tren nyata mamalia jantan untuk menunjukkan telomer yang lebih pendek (Barrett dan Richardson 2011 ), yang dapat dibalik pada beberapa spesies. Jantan heterogametik mungkin dirugikan jika mutasi yang merusak memengaruhi gen terkait-X yang terlibat dalam pemeliharaan telomer, seperti DKC1 pada manusia (Barrett dan Richardson 2011 ). Pola ini mungkin dibalik dalam taksa seperti burung dan lepidoptera, di mana betina adalah jenis kelamin heterogametik (Horn et al. 2011 ). Jantan P. hastatus adalah heterogametik, jadi ini tidak dapat menjelaskan pola yang kami amati, dan prediksi ini juga tidak didukung oleh meta-analisis perbedaan jenis kelamin dalam panjang telomer (Remot et al. 2020 ). Atau, variasi dalam panjang telomer dapat muncul sebagai akibat dari dimorfisme ukuran seksual. Di antara mamalia yang menunjukkan dimorfisme ukuran seksual, jantan cenderung menjadi jenis kelamin yang lebih besar, tetapi ini tidak selalu terjadi (Tombak et al. 2024 ). Investasi dalam pertumbuhan dan pemeliharaan massa tubuh yang lebih besar dikaitkan dengan peningkatan replikasi sel yang, jika tidak ada aktivitas telomerase, diharapkan dapat mengurangi panjang telomer (Barrett dan Richardson 2011 ). P. hastatus jantan lebih besar baik dalam ukuran rangka maupun massa tubuh tetapi memiliki telomer yang lebih panjang, jadi prediksi ini juga tidak menjelaskan temuan kami.
Pengamatan kami bahwa P. hastatus jantan cenderung memiliki telomer yang lebih panjang, meskipun ukuran tubuh lebih besar dan kematian lebih awal, bertentangan dengan prediksi kami. Perbedaan jenis kelamin dalam panjang telomer dapat bersifat konstitutif atau mungkin muncul jika jenis kelamin berinvestasi secara berbeda dalam pemeliharaan telomer, misalnya, melalui enzim telomerase, yang baru-baru ini ditemukan diekspresikan dalam jaringan sayap kelelawar (Athar et al. 2024 ; Li et al. 2023 ; Power et al. 2023 ) dan ekspresinya berkorelasi dengan massa di seluruh mamalia (Gomes et al. 2011 ). Temuan kami tentang telomer yang lebih panjang pada jantan kelelawar dengan mortalitas yang cenderung ke jantan tampak konsisten dengan pandangan bahwa pola telomer yang lebih pendek pada spesies yang berumur lebih panjang (Gomes et al. 2011 ), mungkin karena peran degradasi telomer dalam pertahanan tumor, mungkin terulang pada tingkat dalam spesies (Tissier et al. 2022 ). Jantan P. hastatus yang berumur relatif pendek mungkin kurang terpapar pada seleksi pada pertahanan tumor karena mortalitas yang lebih awal, yang dapat mendukung seleksi terhadap efek buruk telomer yang lebih pendek di awal kehidupan (Wilbourn et al. 2018 ). Di sisi lain, hubungan terbalik antara massa spesies dan panjang telomer (Pepke dan Eisenberg 2022 ) akan, jika diterapkan pada tingkat dalam spesies, memprediksi bahwa jantan yang lebih besar akan memiliki telomer yang lebih pendek. Dalam kasus P. hastatus , efek seperti itu mungkin dinetralisir oleh mortalitas jantan yang sangat dipercepat. Namun, pada titik ini, pengamatan kami juga konsisten dengan temuan meta-analisis terkini yang menunjukkan bahwa di antara vertebrata dengan dimorfisme ukuran seksual yang cenderung ke arah jantan, jantan cenderung memiliki telomer yang lebih panjang (Remot et al. 2020 ). Remot et al. tampaknya tidak yakin dengan temuan ini dan menemukan bahwa penghapusan kumpulan data mandrill ( Mandrillus sphinx ) (Beaulieu et al. 2017 ) membuat pola ini tidak signifikan, tetapi kami pikir hal ini mungkin perlu dipertimbangkan lebih lanjut.
Kami mencatat peringatan tertentu yang perlu dipertimbangkan ketika menafsirkan hasil kami. Seperti dalam studi cross-sectional lainnya, ada kemungkinan bahwa pola yang kami amati dipengaruhi oleh hilangnya individu dengan telomer yang lebih pendek secara selektif. Selain itu, seleksi pada panjang telomer dapat berbeda antara jenis kelamin (Bauch et al. 2020 ), dan karena itu dapat berkontribusi pada perbedaan yang diamati dalam panjang telomer baik di seluruh usia maupun antara jenis kelamin. Misalnya, hilangnya kelelawar dengan telomer yang lebih pendek secara selektif karena bias jantan, mungkin terkait dengan sistem perkawinan yang sangat poligini, dapat menyembunyikan pola pengurangan telomer pada jantan dari sampel cross-sectional kami dan menunjukkan bahwa jantan cenderung memiliki telomer yang lebih panjang di antara individu yang seusia. Selanjutnya, distribusi usia dalam sampel kami berbeda antara jenis kelamin. Sehubungan dengan rentang usia, ini mewakili secara biologis (Adams et al. 2025 ; Wilkinson et al. 2024 ), dan kami menemukan bahwa membatasi analisis kami pada subsampel yang tidak bias tidak memengaruhi interpretasi telomer yang lebih panjang pada pria. Akhirnya, kami menemukan bahwa pengukuran panjang telomer dipengaruhi oleh durasi penyimpanan sampel. Ini tidak terduga, karena penelitian sebelumnya yang menggunakan qPCR untuk mengukur telomer kelelawar tidak mengamati efek durasi penyimpanan dalam kondisi serupa selama durasi yang lebih lama (Power et al. 2023 ), tetapi penyertaan faktor-faktor ini dalam model kami harus memperhitungkan variasi teknis ini.
Mengingat minat yang luas pada faktor-faktor yang mendasari umur panjang yang panjang pada kelelawar (Gorbunova et al. 2020 ; Teeling 2021 ), hanya ada sedikit studi tentang panjang telomer. Di antara beberapa studi yang diterbitkan, ada bukti beragam tentang penurunan terkait usia di seluruh taksa (Foley et al. 2018 , 2020 ; Forest 2022 ; Ineson et al. 2020 ; Power et al. 2023 ). Studi kami menambah keragaman filogenetik yang terwakili untuk klade ini, sebagai satu-satunya perwakilan Phyllostomidae , salah satu kelompok mamalia paling beragam secara ekologis (Leiser-Miller dan Santana 2020 ). Pola pengurangan telomer terkait usia bervariasi di antara spesies kelelawar, dengan spesies genus Myotis tampaknya tidak menunjukkan bukti pengurangan (Foley et al. 2018 ; Ineson et al. 2020 ). Namun, dengan mempertimbangkan temuan kami, tampaknya ada tren umum penurunan panjang telomer terkait usia pada spesies kelelawar (Foley et al. 2018 ; Forest 2022 ; Power et al. 2023 ), yang menunjukkan bahwa pemeliharaan telomer bukanlah ciri pembeda yang mendasari umur panjang mereka yang luar biasa.