
Bagaimana osilasi iklim yang cepat dan luas pada akhir Pleistosen memengaruhi perluasan populasi di seluruh spesies yang secara ekologis berbeda? Dalam artikel From the Cover dalam terbitan Molecular Ecology ini , Harrington dkk. ( 2024 ) merekonstruksi sejarah demografi sembilan spesies ular atau kompleks spesies dari Nearctic Timur untuk menjawab pertanyaan ini. Dengan memanfaatkan data genomik yang baru dihasilkan dan dipublikasikan, penulis menguji hipotesis bahwa garis keturunan ular ini menunjukkan perluasan demografi yang sinkron setelah Maksimum Glasial Terakhir (LGM). Temuan mereka menunjukkan bahwa sebagian besar garis keturunan ular yang diuji memang menunjukkan bukti perluasan populasi yang sinkron setelah Pleistosen, meskipun kurang terkait erat dengan pemanasan pasca-LGM daripada yang diharapkan. Sementara beberapa garis keturunan menunjukkan pertumbuhan eksponensial setelah LGM, garis keturunan lainnya berkembang lebih bertahap sepanjang Pleistosen. Untuk menjawab pertanyaan luas tentang apa yang mungkin menjelaskan variasi dalam respons perluasan di seluruh spesies, penulis juga melakukan analisis korelasi antara faktor lingkungan dan jarak genetik dalam spesies, dan menyimpulkan bahwa sifat ekologis khusus spesies kemungkinan menjelaskan pola demografi yang berbeda di seluruh spesies. Kesamaan yang luas dalam waktu perluasan di antara spesies memberikan bukti baru yang meyakinkan untuk efek konsisten dari pemanasan pasca-Pleistosen pada populasi ektoterm, seperti ular. Secara umum, studi ini semakin memperkuat Pleistosen akhir sebagai model yang berharga untuk memahami respons khusus spesies terhadap perubahan iklim masa lalu, dan potensi pemahaman tersebut untuk memprediksi bagaimana spesies dapat merespons secara berbeda terhadap perubahan iklim di masa mendatang.
Transisi antara periode glasial dan interglasial selama Pleistosen (2 juta tahun terakhir), khususnya pemanasan cepat setelah Maksimum Glasial Terakhir (LGM) sekitar 18.000 tahun yang lalu, merupakan pergeseran iklim paling signifikan dalam sejarah Bumi terkini (Clark et al. 2009 ). Dengan demikian, pergeseran iklim ini berdampak besar pada distribusi flora dan fauna, yang menyebabkan pergeseran yang sesuai dalam distribusi dan ukuran populasi spesies (Hewitt 2000 ), khususnya yang terdistribusi di Nearctic. Pergeseran skala besar ini didorong oleh banyak faktor yang saling berhubungan dan tidak saling eksklusif, termasuk: (a) mundurnya lapisan es besar yang pernah menyelimuti Amerika Utara bagian utara (Gambar 1 ), (b) kenaikan suhu yang signifikan—hingga 9°C di beberapa wilayah (Gambar 2 ; Annan dan Hargreaves 2013 )—disertai dengan kondisi yang lebih kering, (c) munculnya rezim iklim baru yang tidak ada selama periode glasial, dan (d) perombakan ekologi, yang memaparkan spesies terhadap tekanan selektif baru yang terkait dengan ketersediaan makanan, predasi, dan kompetisi (Jackson dan Overpeck 2000 ).
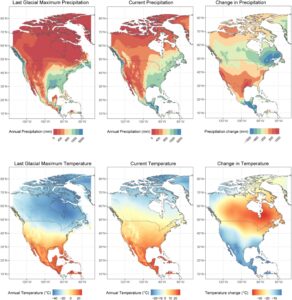
Memahami bagaimana spesies merespons peristiwa iklim ekstrem di masa lalu ini dapat memberikan wawasan berharga tentang bagaimana spesies yang berbeda dapat merespons perubahan iklim di masa mendatang. Memang, banyak penelitian telah memeriksa persamaan dan perbedaan dalam respons lintas spesies terhadap pemanasan pasca-LGM (misalnya, Jackson dan Overpeck 2000 ) dan menemukan pola yang sangat berbeda lintas spesies dalam respons mereka. Banyak spesies menyesuaikan diri dengan perubahan iklim dengan menggeser jangkauan mereka ke garis lintang atau ketinggian yang lebih tinggi untuk melacak, dan dengan demikian mempertahankan ceruk iklim mereka (Chen et al. 2011 ). Spesies lain tampaknya telah bertahan dalam rentang historis mereka dengan memperluas ceruk mereka untuk mentolerir kondisi yang lebih hangat (Jezkova et al. 2011 ), sementara yang lain menjajah wilayah yang sama sekali baru dengan iklim non-analog, seringkali jauh melampaui distribusi mereka sebelumnya (misalnya, Jezkova et al. 2016 ; Svenning dan Skov 2007 ).
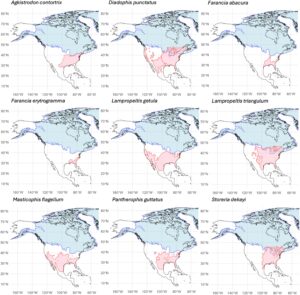
Dengan memeriksa 17 garis keturunan berbeda di sembilan spesies ular atau kompleks spesies (Gambar 1 ), Harrington et al. ( 2024 ) menyelidiki dampak perubahan iklim Pleistosen akhir pada pergeseran ukuran populasi ular. Berdasarkan kesimpulan dari data genomik, mereka menguji prediksi luas bahwa garis keturunan ular Nearctic ini menunjukkan bukti ekspansi demografi sinkron yang bertepatan dengan akhir Pleistosen. Landasan logis untuk ekspektasi ini didasarkan pada prediksi bahwa mundurnya lapisan es dan iklim yang memanas yang terkait dengan Maksimum Glasial Terakhir (LGM) seharusnya menyebabkan peningkatan ketersediaan habitat bagi organisme ektotermik ini, yang distribusinya sering dibatasi oleh suhu lingkungan. Para penulis menemukan dukungan untuk ekspansi populasi di semua kecuali tiga garis keturunan ular. Namun, mereka juga menemukan bahwa tingkat dan waktu ekspansi bervariasi di antara 17 garis keturunan, dan dalam banyak garis keturunan, ekspansi dimulai jauh sebelum Maksimum Glasial Terakhir (LGM), dalam beberapa kasus sedini 2 juta tahun sebelum sekarang. Para penulis menyarankan bahwa variasi dalam waktu perluasan populasi yang diamati di seluruh garis keturunan didorong oleh respons khusus spesies terhadap variabel iklim dan lingkungan. Namun, berdasarkan analisis tentang bagaimana kondisi lingkungan saat ini memengaruhi jarak genetik di seluruh populasi dalam garis keturunan yang berbeda, mereka tidak menemukan tren yang jelas yang menjelaskan mengapa garis keturunan yang berbeda menunjukkan riwayat perluasan yang berbeda. Meskipun tidak diuji secara langsung dalam makalah tersebut, hasil mereka tampaknya menunjukkan tren umum di mana garis keturunan utara dan timur tampaknya menunjukkan perluasan yang lebih besar dan lebih selaras dengan LGM dibandingkan dengan garis keturunan selatan dan barat, kemungkinan karena mundurnya lapisan es, yang membuka habitat baru yang bebas persaingan. Perluasan garis keturunan selatan dan barat mungkin didorong oleh suhu yang lebih hangat secara keseluruhan. Selain itu, ular berbadan besar tampaknya menunjukkan perluasan yang lebih erat kaitannya dengan waktu LGM, mungkin karena kemampuan penyebarannya yang lebih tinggi dan kebutuhan makanan yang lebih luas. Sementara para penulis berhenti menawarkan interpretasi khusus untuk variasi dalam riwayat perluasan yang mereka amati di seluruh spesies, ukuran sampel yang relatif kecil membatasi potensi untuk analisis yang lebih luas terhadap faktor-faktor yang berkovariasi dengan pola perluasan.
Studi ini menyoroti nilai ektotermik sebagai sistem model untuk mempelajari konsekuensi ekologis dan evolusi dari perubahan lingkungan, mengingat ketergantungan fisiologisnya yang kuat pada suhu lingkungan. Mempertimbangkan sifat ektotermik ular, masuk akal untuk berasumsi bahwa harus ada hubungan yang sederhana dan konsisten antara pemanasan iklim dan perluasan populasi. Sementara temuan Harrington et al. menunjukkan tren ini pada skala kasar, temuan tersebut juga memberikan bukti adanya variasi yang agak mengejutkan tinggi dalam respons terhadap pemanasan pasca-LGM di seluruh spesies ular. Temuan ini menimbulkan pertanyaan luas tentang bagaimana garis keturunan ular yang berbeda mungkin telah terdampak oleh perubahan iklim masa lalu dengan cara yang sangat berbeda, dan bagaimana mereka mungkin terdampak secara berbeda oleh perubahan iklim di masa mendatang. Temuan bahwa banyak garis keturunan tampaknya telah mengalami perluasan yang relatif bertahap selama periode Pleistosen yang panjang—sebelum Maksimum Glasial Terakhir (LGM)—mengejutkan dan menantang ekspektasi langsung bahwa suhu yang lebih hangat secara langsung menyebabkan perluasan populasi pada ektotermik. Sebaliknya, hal itu mungkin menunjukkan bahwa perubahan iklim itu sendiri, dan mungkin gangguan habitat yang terkait dengan perubahan iklim, menciptakan peluang utama yang memfasilitasi perluasan garis keturunan tertentu. Secara keseluruhan, temuan mereka menyoroti kompleksitas variasi dalam respons terhadap perubahan iklim di seluruh spesies ular, dan membuka pertanyaan baru tentang kompleksitas hubungan iklim-ekologi yang mungkin mendasari perbedaan khusus spesies ini.
Selain ular, karya Harrington dkk. memberikan contoh menarik yang menggambarkan bagaimana studi tentang respons spesies terhadap peristiwa pemanasan di masa lalu dapat memberikan wawasan utama tentang bagaimana organisme dapat merespons pemanasan iklim di masa mendatang. Temuan mereka lebih jauh menyoroti pentingnya bergerak melampaui model sederhana yang mengasumsikan pemanasan saja yang mendorong perluasan atau penyusutan populasi, dan sebaliknya memotivasi pertimbangan tentang interaksi multidimensi yang kompleks antara iklim, bentang alam, dan biologi khusus spesies yang mungkin lebih realistis menggambarkan kemungkinan respons spesies terhadap perubahan iklim. Langkah alami selanjutnya adalah membangun pendekatan ini dan pendekatan serupa dengan menyelidiki bagaimana perluasan jangkauan berhubungan dengan sifat khusus spesies, menggunakan peristiwa iklim di masa lalu dan kumpulan spesies komparatif yang lebih besar sebagai eksperimen alami untuk menguji hipotesis ini lebih lanjut.
Kemajuan dalam metode untuk menyimpulkan sejarah demografi garis keturunan, bersama dengan biaya pengumpulan data genom yang menurun dengan cepat, sekarang membuatnya semakin memungkinkan untuk menskalakan pertanyaan yang terkait dengan respons skala komunitas terhadap perubahan iklim ke tingkat baru yang menggabungkan koleksi spesies komparatif yang semakin besar dan beragam. Sementara studi Harrington et al. bergantung pada penggunaan sekuensing genom representasi-tereduksi (yaitu, sekuensing RAD) dan Spektrum Frekuensi Situs (SFS) untuk memperkirakan ukuran populasi dari waktu ke waktu, pendekatan lain yang lebih kuat menggunakan data sekuensing ulang genom secara keseluruhan juga dapat dimanfaatkan untuk membuat inferensi demografi resolusi yang lebih tinggi. Misalnya, potensi untuk menerapkan model Sequentially Markovian Coalescent (SMC) ke sejumlah kecil genom individu, atau bahkan satu individu, untuk menyimpulkan sejarah demografi, secara besar-besaran meningkatkan potensi cakupan dan skala keanekaragaman yang dapat ditangani oleh studi komparatif tersebut. Ke depannya, ada potensi yang luas dan belum dimanfaatkan untuk memperluas studi perbandingan yang menyimpulkan demografi historis dari sekumpulan besar spesies, atau bahkan seluruh komunitas, untuk merekonstruksi respons masa lalu terhadap perubahan iklim, dan dengan demikian memperkirakan variasi dalam respons iklim. Pendekatan semacam itu dapat memberikan wawasan baru yang mendasar tentang potensi ketahanan spesies terhadap perubahan iklim dan membantu memahami bagaimana variasi dalam respons khusus spesies terhadap perubahan iklim dapat terwujud pada skala ekologi secara keseluruhan.