
Abstrak
- Respons ektotermik terhadap suhu permisif yang mendukung pertumbuhan, pemeliharaan, dan perbaikan biasanya dimodelkan oleh kurva kinerja termal (TPC). Sebaliknya, model waktu kematian termal (TDT) sangat cocok untuk menggambarkan akumulasi cedera eksponensial yang kuat yang terjadi pada suhu yang penuh tekanan. Meskipun setiap model dipahami dengan baik, kompleksitas muncul ketika batas model dilintasi.
- Dengan menggunakan larva lalat tentara hitam ( Hermetia illucens ), kami mengkarakterisasi kurva kinerja termal untuk pertumbuhan. Kami kemudian menerapkan suhu optimal (TPC opt = 35,6°C) sebagai titik pemulihan untuk menilai gangguan pertumbuhan pada larva setelah paparan stres termal yang terkontrol dan spesifik dosis di luar batas atas TPC (TPC maks ~ 42°C).
- Di atas 42°C, cedera terakumulasi secara aditif menuju dosis mematikan, dengan intensitas yang lebih tinggi mempercepat akumulasi cedera dan memperpendek waktu bertahan hidup (Lt50 ) , seperti yang diprediksi oleh model TDT. Menggunakan parameter TDT, kami menerapkan dosis tetap stres termal (misalnya 10%, 20%, 30% dari dosis mematikan) untuk menunjukkan hubungan linear negatif antara dosis stres dan pengurangan berikutnya dalam laju pertumbuhan spesifik ketika larva dikembalikan ke suhu permisif. Secara khusus, cedera panas subletal yang sesuai dengan 1% dari Lt50 mengurangi laju pertumbuhan relatif sebesar 1,41% hari -1 ketika laju pertumbuhan dievaluasi setelah 1 hari. Seiring waktu, larva yang terluka pulih, dan gangguan pertumbuhan yang tampak, oleh karena itu, berkurang ketika dievaluasi setelah 7 hari (0,14% hari -1 ) dan 10 hari (0,09% hari -1 ). Sebaliknya, pengurangan awal dalam pertumbuhan secara bertahap terwujud sebagai perkembangan yang tertunda, dengan larva mencapai tahap prapupa 0,0247 hari kemudian untuk setiap 1% dosis mematikan.
- Secara kolektif, temuan-temuan ini menunjukkan bagaimana pendekatan yang konsekuen dan berbasis model dapat mengukur biaya stres panas subletal pada hewan yang mengalami fluktuasi suhu antara kisaran yang memungkinkan dan yang penuh tekanan. Kami menunjukkan bahwa biaya stres subletal memiliki konsekuensi kebugaran yang bertahan lama, meningkat secara linear dengan dosis stres dan bersifat aditif di seluruh kisaran suhu yang penuh tekanan. Mengakui dan memparameterisasi efek-efek ini sangat penting untuk memperkirakan dampak perubahan iklim dalam sistem alami dan mengukur biaya stres panas dalam produksi serangga skala besar.
Baca Ringkasan Bahasa Sederhana gratis untuk artikel ini di blog Jurnal
1. PENDAHULUAN
Terjadinya peristiwa panas ekstrem meningkat seiring meningkatnya suhu global dengan perubahan iklim antropogenik (IPCC, 2021 ; Perkins-Kirkpatrick & Lewis, 2020 ). Dengan demikian, telah menjadi tujuan utama dalam ekologi dan konservasi untuk memahami bagaimana pemanasan berdampak pada hewan ektotermik (Deutsch et al., 2008 ; Filazzola et al., 2021 ; González-Tokman et al., 2020 ; Harvey et al., 2023 ; Jørgensen et al., 2022 ). Batas toleransi termal yang diukur saat hewan terpapar suhu yang menegangkan dan merugikan sering kali diusulkan sebagai prediktor yang berharga dari distribusi biogeografis ektoterm, termasuk serangga (Bennett et al., 2021 ; Kellermann et al., 2012 ; Sunday et al., 2012 , 2019 ). Namun, perdebatan yang cukup besar tetap ada mengenai metrik toleransi termal mana yang paling baik mencirikan distribusi spesies global (Parratt et al., 2021 ) dan ciri-ciri riwayat hidup, seperti kemampuan reproduksi, pertumbuhan, dan perkembangan semakin banyak digunakan dalam model distribusi spesies (Deutsch et al., 2008 ; Ørsted et al., 2024 ; Overgaard et al., 2014 ; Parratt et al., 2021 ; Piyaphongkul et al., 2012 ; Walsh et al., 2021 ). Stres panas subletal terjadi ketika hewan bertahan hidup dari paparan suhu yang menegangkan yang melebihi suhu yang diperlukan untuk mempertahankan homeostasis. Namun, masih kurang dipahami bagaimana hewan ektotermik merespons fluktuasi antara suhu yang memungkinkan dan suhu yang penuh tekanan (Colinet et al., 2015 ; Jørgensen et al., 2022 ; Nedvěd et al., 1998 ; Ørsted et al., 2022 ).
Biologi termal ektoterm umumnya dievaluasi baik dalam rentang suhu permisif, yang mencakup suhu optimal dan suboptimal yang mendukung pertumbuhan dan reproduksi (Kingsolver et al., 2023 ; Sinclair et al., 2016 ; Vasseur et al., 2014 ), atau dalam rentang stres, di mana stres panas menyebabkan akumulasi cedera yang akhirnya menyebabkan kematian (Fry, 1946 , 1971 ; Jørgensen et al., 2022 ). Rentang suhu permisif beroperasi dengan kepekaan termal dan proses biologis yang secara fundamental berbeda dari yang mengatur (hilangnya) kinerja pada suhu stres (Jørgensen et al., 2022 ; Ørsted et al., 2022 ; Gambar S1 ). Untuk memformalkan pemahaman ini, Ørsted et al. ( 2022 ) mengusulkan penggunaan suhu kritis ( T C ) untuk menggambarkan batas antara rentang suhu permisif dan stres, berdasarkan keseimbangan antara proses homeostatis dan disruptif, masing-masing. Sementara T C secara konseptual dianggap sebagai suhu spesifik, itu lebih akurat sebagai rentang suhu yang sempit. Di bawah T C , efek suhu pada sifat kebugaran seperti pertumbuhan dan reproduksi dijelaskan dengan baik dengan kurva kinerja termal (TPC), yang non-linier dan biasanya menunjukkan pola asimetris dan miring ke kiri (Cossins & Bowler, 1987 ; Sinclair et al., 2016 ). Saat suhu meningkat, kinerja organisme meningkat secara eksponensial dari ambang batas minimum yang diizinkan (TPC min ), mencapai puncaknya pada suhu optimal (TPC opt ) dan menurun tajam, mendekati nol saat suhu mendekati batas termal atas untuk kinerja (TPC max ) (Huey et al., 2012 ; Huey & Stevenson, 1979 ; Sinclair et al., 2016 ). Sementara TPC max untuk sifat kronis seperti pertumbuhan dan reproduksi menunjukkan suhu saat akumulasi energi bersih mendekati nol, itu bukan perkiraan langsung dari suhu mematikan (CT max ) (Ørsted et al., 2022 ; lihat Lampiran S1 ).
Di luar TPC maks , dan di atas T C , efek negatif dari suhu yang penuh tekanan mendominasi dan ini dijelaskan dengan baik oleh model waktu kematian termal (TDT), di mana waktu toleransi menurun dengan cepat dan eksponensial saat intensitas tekanan panas (yaitu suhu) meningkat (Cossins & Bowler, 1987 ; Jørgensen et al., 2019 ; Rezende et al., 2014 ). Dalam rentang suhu ini, cedera panas terakumulasi secara aditif dan jika tekanan panas yang cukup terakumulasi, itu dapat menyebabkan keruntuhan dan kematian organisme. Bukti menunjukkan bahwa proses biologis yang menyebabkan cedera panas di seluruh rentang suhu yang penuh tekanan serupa, apakah tekanan panas itu intens atau sedang (Fry, 1946 ; Jørgensen et al., 2021 ; Ørsted et al., 2022 ). Suhu tinggi dalam kisaran yang menimbulkan stres menyebabkan kematian lebih cepat karena stres terakumulasi lebih cepat, memperpendek waktu untuk mencapai dosis yang mematikan (misalnya intensitas tinggi menghasilkan waktu bertahan hidup yang lebih pendek, dan sebaliknya). Konsisten dengan pandangan ini, adalah mungkin untuk mengekspos hewan pada dosis spesifik cedera panas subletal yang sesuai dengan sebagian kecil (misalnya, 10% atau 20%) dari dosis yang mematikan.
Fluktuasi suhu yang terjadi secara eksklusif di bawah atau di atas T C (masing-masing secara eksklusif dalam rentang permisif atau stres) dipahami dengan baik dan dijelaskan menggunakan model TPC atau TDT. Namun, kompleksitas meningkat ketika suhu berfluktuasi melintasi T C , antara rentang permisif dan stres. Studi terbaru dan historis menunjukkan bahwa stres panas (dan dingin) subletal dapat dikurangi dan diperbaiki ketika paparan stres terganggu oleh fluktuasi dalam rentang suhu permisif (Colinet et al., 2015 ; Nedvěd et al., 1998 ; Ørsted et al., 2022 ). Dalam studi ini, kami membalikkan perspektif ini untuk memeriksa biaya dan keterbatasan yang dikenakan pada proses biologis permisif pada hewan setelah paparan dosis stres panas subletal yang terpisah. Melalui eksperimen kami, kami bertujuan untuk membangun kerangka kerja untuk mengukur dampak/biaya langsung dari cedera panas subletal pada hewan ektotermik. Biaya ini kemungkinan terjadi di lingkungan alami dan sistem produksi serangga, yang berdampak pada kebugaran, pertumbuhan dan kemungkinan dinamika populasi atau produktivitas.
Untuk menyelidiki biaya stres panas subletal, kami menggunakan larva lalat tentara hitam (BSFL) ( Hermetia illucens ); spesies yang tumbuh cepat dan mudah diakses dengan toleransi suhu yang luas (~12°C–42°C) (Chia et al., 2018 ; Siddiqui et al., 2024 ). Lalat Tentara Hitam yang aslinya berasal dari Amerika kini tumbuh subur di wilayah tropis dan beriklim sedang (Wang & Shelomi, 2017 ). BSFL digunakan secara global dalam industri karena kemampuannya yang efisien dalam meningkatkan sumber daya berkualitas rendah menjadi protein dan lemak (Abd El-Hack et al., 2020 ; Siddiqui et al., 2024 ). Sistem produksi intensitas tinggi untuk BSFL dan sistem produksi ektotermik lainnya (misalnya akuakultur, budidaya serangga) sering beroperasi dalam kondisi termal yang tinggi untuk mengoptimalkan laju pertumbuhan, tetapi berisiko menimbulkan stres (Islam et al., 2022 ; Schøn et al., 2024 ). Untuk tujuan penelitian ini, pertama-tama kami mengkarakterisasi kurva kinerja termal (TPC) untuk pertumbuhan BSFL untuk mengidentifikasi suhu yang mendukung pertumbuhan secara optimal (TPC opt ) dan untuk menilai batas atas kisaran suhu permisif yang mendukung pertumbuhan bersih (TPC max ). Selanjutnya, kami menilai sensitivitas stres panas di berbagai suhu stres menggunakan model waktu kematian termal (TDT) yang menggambarkan bagaimana cedera (misalnya kebugaran negatif) terakumulasi pada tingkat yang meningkat dengan meningkatnya intensitas stres (suhu) (Lampiran S1 ). Menggabungkan model-model ini di seluruh rentang permisif dan stres memungkinkan kita untuk mengukur biaya jangka pendek dari stres panas subletal dengan mengukur gangguan pertumbuhan pada larva ketika mereka dikembalikan ke kondisi permisif setelah dosis stres yang berbeda dan terdefinisi dengan baik. Kami meneliti (a) apakah biaya dalam hal pertumbuhan yang berkurang meningkat secara linear dengan dosis stres fraksional; (b) apakah biaya stres panas pada pertumbuhan secara kuantitatif serupa, terlepas dari intensitas stres panas (suhu) yang diterapkan untuk mencapai dosis stres tertentu; (c) apakah biaya stres panas dari dua intensitas yang berbeda bersifat aditif; (d) apakah gangguan pertumbuhan jangka pendek dikompensasi dari waktu ke waktu melalui waktu perkembangan yang diperpanjang. Dengan menggunakan data ini, kami kemudian membahas dan mengembangkan kerangka kuantitatif untuk mengevaluasi dampak paparan suhu yang berfluktuasi antara rentang yang memungkinkan dan yang penuh tekanan, dengan implikasi untuk memahami respons termal di ekosistem alami dan lingkungan produksi.
2 BAHAN DAN METODE
2.1 Hewan percobaan dan pakan
Larva lalat tentara hitam ( H. illucens ) diperoleh sebagai larva instar pertama dari Enorm Biofactory A/S (Flemming, Denmark) dan dibesarkan hingga ukuran 2–5 mg (setara dengan instar ke-4-5 (Gligorescu et al., 2019 )) sebelum digunakan dalam percobaan. Kecuali dinyatakan lain, larva dipelihara pada kelembapan tinggi dalam lemari yang suhunya diatur pada 35°C. Larva dibesarkan dengan diet percobaan yang terdiri dari campuran 2:1 air dan ‘paco chicken feed start’ kering, yang bersumber dari Land&Fritid (Ringsted, Denmark). Pakan tersebut mengandung kedelai, oat, jagung, gandum, dan barley dengan komposisi nutrisi protein kasar 18,5%, lemak kasar 3,3%, lignin 4%, abu kasar 5,5%, lisin 10 g/kg, metionin 4,1 g/kg, sistein 3,2 g/kg, kalsium 10 g/kg, fosfor 5,5 g/kg, natrium 1,6 g/kg, dan seng 105 mg/kg. Rasio air terhadap pakan 2:1 dipilih untuk memastikan konsistensi pakan yang lembap tanpa air bebas, sehingga mencegah larva tenggelam.
2.2 Protokol Eksperimen
Percobaan dirancang untuk mengkarakterisasi sensitivitas termal larva lalat tentara hitam (BSFL) dalam rentang suhu yang memungkinkan dan yang penuh tekanan dan untuk mengevaluasi konsekuensi transisi antara rentang ini. Secara khusus, kami bertujuan untuk (1) Mengkarakterisasi kurva kinerja termal untuk pertumbuhan dalam rentang suhu yang memungkinkan; (2) Melakukan analisis waktu kematian termal untuk memeriksa hubungan antara durasi toleransi dan intensitas panas dalam suhu yang penuh tekanan; (3) Mengevaluasi biaya cedera panas setelah paparan subletal terhadap suhu yang penuh tekanan, berdasarkan gangguan pertumbuhan jangka pendek dan jangka panjang pada larva yang dikembalikan ke suhu yang memungkinkan. Penelitian ini tidak memerlukan persetujuan etis.
Selama pemaparan eksperimental pada suhu tertentu, larva ditempatkan dalam tabung Eppendorf 2 mL yang berisi sedikit pakan ayam yang dipanaskan terlebih dahulu (kecuali dinyatakan lain), dengan tutup berlubang untuk memungkinkan pertukaran gas. Setiap tabung Eppendorf ditempatkan secara individual dalam tabung plastik 50 mL, yang direndam dalam penangas air yang suhunya diatur. Tabung 50 mL berisi 3–5 mL air untuk mempertahankan lingkungan termal yang terkontrol dan lembab sambil memastikan akses udara terus-menerus untuk larva (lihat ilustrasi di Lampiran S2 ). Semua suhu perlakuan dipantau menggunakan termometer referensi (termometer Digital Testo 108) yang ditempatkan dalam tabung Eppendorf dengan pakan untuk mensimulasikan kondisi larva. Pengukuran massa larva dilakukan menggunakan skala 6-desimal (Sartorius Lab Instruments GmbH & Co. KG, Göttingen—MSE6.6S-000-DM).
2.3 Kinerja termal pada suhu yang diizinkan
Untuk menilai kinerja BSFL pada berbagai suhu yang diizinkan, kami mengukur laju pertumbuhan spesifik (SGR) setelah paparan selama 24 jam pada salah satu dari 47 perlakuan suhu konstan yang berkisar dari 12,3°C hingga 42,4°C. Untuk setiap suhu, setidaknya 6 larva digunakan (massa rata-rata: 4,02 mg, SD ± 0,86, n = 561). Setiap larva ditimbang satu per satu, ditempatkan dalam tabung Eppendorf (2 mL) yang berisi 1 g pakan ayam (tidak dipanaskan terlebih dahulu) dan dikenakan pada suhu yang diinginkan. Setelah 24 jam, larva diambil sampelnya dan ditimbang ulang, dan SGR setiap larva dihitung:
![]()
Penanganan larva kecil kadang-kadang menyebabkan cedera, dan beberapa individu terpengaruh secara buruk pada suhu tertinggi yang digunakan untuk menilai laju pertumbuhan. Untuk memastikan bahwa hanya laju pertumbuhan hewan yang layak yang dipertimbangkan, data dari larva yang mati atau tampak terluka dikecualikan. Selain itu, karena beberapa larva kemungkinan bertransisi antara instar selama periode pertumbuhan 24 jam, kami mengamati replikasi di mana pertumbuhannya sangat berbeda dari rata-rata kelompok. Untuk mengatasi hal ini, outlier dengan nilai SGR yang menyimpang lebih dari ±2 SD dari rata-rata dikecualikan dari analisis. Suhu perlakuan dengan kurang dari enam larva, karena kematian atau penghapusan outlier, dikecualikan dari kumpulan data. Yang penting, pengecualian data outlier memiliki efek yang dapat diabaikan pada estimasi suhu optimal (TPC opt ) dan suhu maksimum untuk pertumbuhan (TPC max ) (lihat Lampiran S3 ).
2.4 Analisis waktu kematian termal pada suhu yang membahayakan
Kurva waktu kematian termal (TDT) dievaluasi dengan memaparkan BSFL ke 9 suhu statis dan penuh tekanan mulai dari 42°C hingga 50°C. Untuk setiap suhu, kami menggunakan minimal 13 replikasi di berbagai interval waktu untuk memodelkan kurva kelangsungan hidup logistik. Durasi paparan untuk setiap suhu dipilih untuk mengamankan data sebelum, dekat dan setelah waktu mematikan (Lt50 ) (lihat Lampiran S4 ). Setiap replikasi berisi 5 BSFL, yang memungkinkan kami untuk mengevaluasi kelangsungan hidup proporsional untuk setiap kombinasi durasi-suhu. Dalam percobaan ini, larva diambil langsung dari kondisi pemeliharaannya pada suhu 35°C dan ditempatkan dalam pakan yang dipanaskan terlebih dahulu dalam tabung Eppendorf berlubang yang segera dipindahkan ke bak air yang suhunya diatur. Setelah durasi paparan yang dipilih, larva dikembalikan ke suhu kamar selama 24 jam setelah itu mortalitas dicatat. Pada setiap suhu eksperimen, Lt50 dievaluasi menggunakan regresi logistik, dengan data kelangsungan hidup dimodelkan sebagai respons binomial terhadap suhu. Kurva TDT kemudian dibuat dengan mentransformasikan nilai Lt 50 ke dalam bentuk logaritma dan memplotkannya terhadap suhu eksperimen. Kemiringan regresi dari analisis ini menggambarkan faktor sensitivitas termal, yang biasanya digambarkan sebagai ‘ z ‘,
 2.5 Gangguan pertumbuhan akibat stres panas subletal
2.5 Gangguan pertumbuhan akibat stres panas subletal
Model TDT memungkinkan kami untuk memperkirakan waktu mematikan (Lt50 ) pada suhu tertentu antara 42 dan 50°C dan menghitung durasi paparan yang setara dengan persentase tetap Lt50 ( dosis mematikan) pada suhu yang penuh tekanan apa pun. Dengan menggunakan pendekatan ini, kami menghitung waktu yang diperlukan untuk mengerahkan 0%, 10%, 20%, 30%, 40%, 50%, 60% dan 70% dari dosis mematikan pada tiga suhu penuh tekanan yang berbeda yang sesuai dengan nilai Lt50 dari 20 menit (47,5°C), 80 menit (45,6°C) dan 320 menit (43,8°C) (lihat Lampiran S5 ). Untuk setiap intensitas stres (10%, 20% dll.) dan kombinasi suhu, kami memaparkan 10 larva dan mengukur ‘biaya stres’ sebagai pengurangan SGR dalam 24 jam berikutnya. Secara khusus, larva ditimbang terlebih dahulu dan ditempatkan secara individual selama 1 jam dalam tabung Eppendorf dengan sejumlah kecil pakan dalam lemari yang suhunya diatur (35°C) untuk memulai pertumbuhan. Selanjutnya, larva dipindahkan ke paparan stres yang ditentukan setelah itu dikembalikan ke lemari suhu dan disimpan pada suhu 35°C selama 23 jam. Larva kemudian ditimbang kembali, dan SGR dihitung (Persamaan 1 ) hanya menggunakan waktu pada suhu optimal dalam penyebut Persamaan ( 1 ) (yaitu waktu pada paparan stres dikecualikan dalam perhitungan SGR). Kelompok kontrol (0% cedera) dimasukkan sebagai referensi untuk semua putaran eksperimen di mana larva kontrol menjalani inkubasi awal 1 jam yang sama pada suhu 35°C, ditangani sebentar, dan kemudian dikembalikan ke lemari selama 23 jam tambahan. Karena SGR sedikit bervariasi antara putaran eksperimen, gangguan pertumbuhan dihitung relatif terhadap SGR kontrol yang tidak stres dari putaran yang sama. Jadi, untuk percobaan ini, kami melaporkan laju pertumbuhan relatif, RGR (RGR = SGR perlakuan/SGR kontrol), sebagai fungsi dosis stres.
2.6 Efek aditif dari cedera termal
Untuk mengeksplorasi efek aditif dari cedera panas subletal pada suhu yang berbeda, kami menguji hipotesis bahwa stres panas pada intensitas tinggi atau rendah bersifat aditif. Suhu stres intensitas tinggi dan rendah dipilih berdasarkan kurva TDT di mana ‘stres intensitas tinggi’ (T1 = 45,6 °C) memiliki Lt 50 yang diharapkan 80 menit, dan suhu ‘stres intensitas rendah’ (T2 = 43,8 °C) memiliki Lt 50 yang diharapkan = 320 menit. Awalnya, larva ( n = 20 untuk setiap kelompok perlakuan) ditempatkan secara individual dalam tabung Eppendorf yang berisi pakan selama 1 jam untuk memulai pertumbuhan. Selanjutnya, larva dipindahkan ke pakan yang dipanaskan terlebih dahulu pada T1 selama durasi yang setara dengan 25% dari Lt 50 setelah itu mereka dipindahkan ke T2 untuk menerima 25% dari Lt 50 pada perlakuan suhu ini (yaitu secara total larva menerima stres suhu yang setara dengan 50% dari Lt 50 ). Kelompok lain menerima perlakuan yang sama, tetapi dalam urutan terbalik (T2 sebelum T1). Terakhir, dua kelompok kontrol menerima paparan stres yang sesuai dengan 50% Lt 50 pada T1 atau T2. Setelah perlakuan, larva dikembalikan ke lemari bersuhu 35°C selama 23 jam, setelah itu RGR dihitung.
2.7 Dampak berkelanjutan dari stres panas sub-mematikan
Untuk mengeksplorasi konsekuensi jangka panjang dari stres panas pada pertumbuhan dalam ‘lingkungan alami’, sejumlah besar BSFL mengalami cedera panas subletal sebelum ditempatkan di lingkungan yang kompetitif dan padat penduduk. Secara khusus, kelompok yang terdiri dari 100 larva dihitung dan ditimbang (massa rata-rata per ulangan: 295 mg untuk 100 larva) sebelum mereka terpapar pada salah satu dari empat perlakuan (masing-masing dengan enam ulangan) dengan dosis stres yang sesuai dengan 0% (kontrol—0 menit), 10% (8 menit), 30% (24 menit) dan 50% (40 menit) dari dosis letal pada suhu 45,6°C (Lt 50 = 80 menit). Selama paparan stres panas, kelompok yang terdiri dari 100 larva ditempatkan dalam tabung 50 mL dengan pakan yang dipanaskan terlebih dahulu dan direndam dalam bak air. Setelah mengalami stres panas, setiap ulangan dipindahkan ke 100 g pakan dan ditempatkan dalam lemari yang suhunya diatur pada 35°C selama 7 hari, setelah itu mortalitas dan pertumbuhan dinilai. RGR dinilai, menggunakan 7 hari sebagai penyebut Persamaan ( 1 ).
Untuk menyelidiki konsekuensi jangka panjang dari stres panas subletal secara lebih rinci, kami memaparkan larva individu pada intensitas stres yang berbeda dan memantau pertumbuhan, waktu perkembangan, dan mortalitas berikutnya selama 12 hari (sampai semua larva berada dalam tahap prapupa). Secara khusus, larva individu ( n = 60 per perlakuan) terpapar cedera subletal yang sesuai dengan 0% (kontrol), 10%, 30% dan 50% dari Lt 50 pada 45,6 ° C (lihat di atas). Setelah paparan stres, larva secara individual ditempatkan pada suhu 35 ° C dalam tabung plastik 50 mL yang berisi pakan ad libitum, dan setiap larva ditimbang setiap hari selama 12 hari. Perkembangan harian dalam massa dievaluasi, dan RGR dinilai pada hari ke-10. Kecepatan perkembangan untuk setiap larva dicatat sebagai waktu hingga massa maksimal, dengan asumsi larva memasuki tahap prapupa non-makan setelahnya. Perhatikan bahwa waktu perkembangan ini menggambarkan perkembangan dari larva ~5 mg (instar ke-4–5) ke tahap prapupa yang tidak makan (instar ke-7). Pada hari ke-12, ada 5 dari 240 larva yang belum mencapai massa maksimal, dan oleh karena itu, kami mengikuti larva ini selama satu hari lagi dan mencatat waktu perkembangan untuk 5 larva ini selama 13 hari. Kombinasi penanganan harian dan paparan stres menyebabkan beberapa kematian dalam percobaan ini, dan oleh karena itu, kami membatasi analisis untuk hanya menyertakan pertumbuhan dan perkembangan larva yang bertahan hidup hingga hari ke-12. Namun, penyertaan atau pengecualian larva yang mati selama periode ini memiliki dampak yang dapat diabaikan pada hasil (lihat Lampiran S6 ). Karena ukuran sampel yang besar, percobaan ini dilakukan dalam dua putaran (semua perlakuan disertakan dalam keduanya), dan putaran eksperimen dimasukkan sebagai variabel dalam analisis statistik kami.
2.8 Pernyataan replikasi
| Skala inferensi | Skala penerapan faktor minat | Jumlah replikasi pada skala yang sesuai | Tujuan |
|---|---|---|---|
| Kurva kinerja termal : Populasi | Subpopulasi | 6–10 larva per perlakuan suhu
47 suhu |
Untuk menilai bagaimana tingkat pertumbuhan bervariasi di seluruh rentang suhu yang diizinkan (Di bawah T C ). Suhu optimal ( T opt ) dari TPC digunakan sebagai ‘titik pemulihan’ saat mengevaluasi biaya stres panas subletal. |
| Kurva Waktu Kematian Termal : Populasi | Subpopulasi | 5 larva per suhu/durasi pengobatan
9 suhu 13–16 durasi untuk setiap perlakuan suhu |
Untuk menguji hubungan antara durasi toleransi dan intensitas panas dalam kisaran suhu stres (di atas T C ) |
| Pertumbuhan (1 hari setelah stres panas) : Populasi | Individu | 10 larva per perlakuan suhu
Tiga suhu stres masing-masing dengan 4 perawatan (+ kontrol) |
Untuk mengevaluasi biaya jangka pendek dari stres panas dengan mengukur gangguan pertumbuhan relatif setelah paparan sub-mematikan terhadap suhu yang penuh tekanan. |
| Efek Aditif : Populasi | Individu | 20 larva per perawatan
4 perawatan (+ kontrol) |
Untuk memeriksa apakah gangguan pertumbuhan akibat paparan dua suhu sub-mematikan yang berbeda mengakibatkan gangguan pertumbuhan aditif |
| Pertumbuhan (7 hari setelah stres panas) : Populasi | Subpopulasi | 6 kali ulangan sebanyak 100 larva untuk setiap perlakuan suhu
4 perawatan (+ kontrol) |
Untuk mengevaluasi biaya persisten dari tekanan panas setelah paparan sub-mematikan terhadap suhu yang penuh tekanan di lingkungan seperti produksi |
| Waktu pertumbuhan & perkembangan (perkembangan penuh setelah stres panas) : Populasi | Individu | 60 larva per perlakuan suhu
4 perawatan (+ kontrol) |
Untuk mengevaluasi biaya jangka panjang dari cedera panas setelah paparan sub-mematikan terhadap suhu yang penuh tekanan |
2.9 Analisis statistik
Semua analisis statistik dilakukan di RStudio (Posit Team, 2023 ; R Core Team, 2023 ), dengan visualisasi data dicapai menggunakan ggplot2 . Kurva kinerja termal (TPC) dimodelkan menggunakan regresi polinomial derajat ke-4, dan simulasi Monte Carlo (100 iterasi, ~90% subsampling acak) memperkirakan varians dalam TPC opt dan TPC max (lihat Lampiran S7 ). Waktu kematian median (Lt50 ) untuk analisis waktu kematian termal (TDT) diperkirakan menggunakan paket MASS , dengan data kelangsungan hidup dimodelkan oleh Model Linear Umum binomial dan kurva kelangsungan hidup divisualisasikan menggunakan kecocokan logistik (lihat Miller, 2010 ). Estimasi waktu kematian median (Lt50 ) yang digunakan untuk menghasilkan kurva waktu kematian termal dirinci dalam Lampiran S8 . Kecocokan model dinilai dengan pseudo- ![]()
Kurva TDT dibuat dengan mencocokkan regresi linier antara Lt50 yang ditransformasikan secara logaritma dan suhu. Perbandingan kelompok efek stres panas aditif di seluruh suhu dinilai menggunakan ANOVA satu arah (basis R) diikuti oleh uji post-hoc Tukey’s HSD ( paket agricolae ).
Hubungan antara cedera dan variabel respons (massa larva dan waktu perkembangan) dievaluasi menggunakan model regresi linier, sementara ANOVA tipe III menguji interaksi cedera-suhu. Ketika variabilitas karena efek acak (misalnya putaran eksperimen) dideteksi dan dikonfirmasi melalui uji rasio-kemungkinan, kami menggunakan model campuran linier menggunakan ( paket nlme ) untuk memperhitungkan efek acak. Analisis kelangsungan hidup dalam percobaan setelah 12 hari pertumbuhan dilakukan menggunakan paket survival . Distribusi kelangsungan hidup kelompok dibandingkan dengan uji Mantel-Cox (log-rank) menggunakan paket survminer untuk menghasilkan dan memvisualisasikan kurva kelangsungan hidup (lihat Lampiran S9 ). Signifikansi statistik ditetapkan pada p < 0,05. Data tersedia dari Dryad Digital Repository: https://doi.org/10.5061/dryad.3r2280gth (Schow-Madsen et al., 2025 ).
3 HASIL
3.1 Kinerja termal pada suhu yang diizinkan
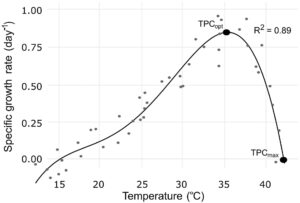
Kurva kinerja termal (TPC) untuk pertumbuhan BSFL dibuat dengan mengukur laju pertumbuhan spesifik (SGR) selama periode 1 hari di 47 suhu statis yang berkisar dari 12,3°C hingga 42,4°C (Gambar 1 ). TPC sangat sesuai dengan polinomial derajat empat ( R 2 = 0,89) yang memperkirakan suhu optimal untuk laju pertumbuhan (TPC opt ) pada 35,6°C ± 0,1°C di mana SGR mencapai puncaknya pada 0,79 hari −1 . Batas atas kisaran suhu yang diizinkan (TPC maks —di mana SGR mencapai 0) diperkirakan pada 42,2°C ± 0,2°C (lihat skrip dan detail analisis Monte Carlo di Lampiran S7 ). Analisis selanjutnya memperlakukan TPC maks (~42°C) sebagai proksi suhu kritis ( T C ), di atas suhu tersebut cedera terjadi dan TPC opt (~35°C) sebagai suhu optimal untuk pertumbuhan, di mana larva dapat pulih untuk mempelajari potensi gangguan pertumbuhan pada larva setelah stres panas subletal.
3.2 Analisis waktu kematian termal—Hubungan antara intensitas tegangan dan waktu toleransi
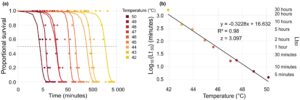
Kami meneliti bagaimana intensitas stres panas (suhu pajanan) memengaruhi durasi toleransi BSFL dengan memperkirakan Lt50 pada 9 suhu stres berbeda yang berkisar dari 42°C hingga 50°C. Pada setiap suhu eksperimen, hubungan antara intensitas dan durasi toleransi sangat sesuai dengan model regresi logistik, dengan nilai pseudo- R2 McFadden berkisar dari 0,61 hingga 0,87 (rata-rata R2 = 0,75). Perkiraan Lt50 bervariasi menurut intensitas stres yang berkisar dari ~1741 menit pada 42°C hingga ~4 menit pada 50°C (Gambar 2a ) (lihat Lampiran S8 ). Dengan menggunakan 9 perkiraan Lt50 , kami kemudian melakukan analisis waktu kematian termal (TDT) (Rezende et al., 2014 ) dengan meregresikan log(Lt50 ) terhadap suhu pajanan. Regresi linier yang dihasilkan memberikan kecocokan yang sangat baik dengan data ( R 2 > 0,98) (Gambar 2b ), yang menunjukkan bagaimana waktu kematian berkurang secara eksponensial dan kuat dengan meningkatnya suhu. Ketika Q 10 dihitung dari kemiringan kurva TDT (Persamaan 2 dan 3 ) kami menemukan sensitivitas termal yang tinggi untuk tingkat cedera panas ( Q 10 = 1690) (Jørgensen et al., 2022 ). Parameter dari analisis TDT juga memungkinkan perhitungan waktu untuk menginduksi proporsi cedera tertentu pada suhu stres tertentu (yaitu pada 45,6 °C, Lt 50 adalah 80 menit, yang berarti bahwa paparan 8, 16 dan 24 menit pada suhu ini sesuai dengan 10%, 20% atau 30% dari Lt 50 , masing-masing (Jørgensen et al., 2021 )).
3.3 Menilai biaya jangka pendek dari stres panas submematikan
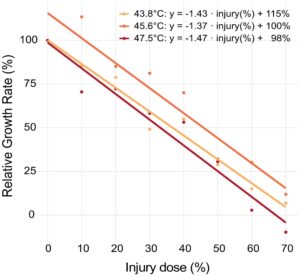
Untuk mengkaji biaya stres panas subletal, kami meneliti gangguan pertumbuhan pada BSFL 24 jam pasca-paparan terhadap berbagai dosis cedera panas. Seperti yang terlihat pada Gambar 3 , laju pertumbuhan relatif (RGR) menurun secara linear dan proporsional seiring dengan peningkatan dosis stres panas. Percobaan ini diulang tiga kali dengan menggunakan intensitas stres yang berbeda untuk menginduksi dosis stres yang diinginkan. Untuk tiga kelompok suhu, kemiringan negatif menunjukkan penurunan RGR yang konsisten dengan peningkatan cedera, dengan pengurangan 1,41% hari -1 untuk setiap peningkatan 1% dalam dosis mematikan (Gambar 3 ). Dengan demikian, larva yang terpapar dosis mematikan 50% kehilangan 69% laju pertumbuhan relatif terhadap kontrol, terlepas dari intensitas stres (suhu stres) yang digunakan untuk menimbulkan dosis stres. Model linear ( R 2 = 0,947) mengungkapkan efek utama yang sangat signifikan dari dosis cedera pada RGR ( p < 0,001 untuk semua kelompok). Tidak ditemukan interaksi signifikan antara cedera dan suhu eksperimen yang digunakan untuk menimbulkan stres (ANOVA Tipe III, p = 0,87), dengan kemiringan serupa di seluruh perawatan (Tukey’s HSD, semua p ≥ 0,86).
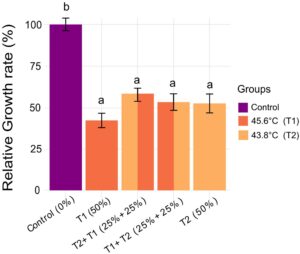
Untuk memeriksa lebih lanjut apakah gangguan pertumbuhan dari intensitas suhu subletal yang berbeda mencerminkan jenis stres panas yang sama, kami menguji apakah menggabungkan cedera dari dua suhu yang penuh tekanan menghasilkan efek aditif pada pertumbuhan. BSFL yang terpapar 50% Lt 50 pada dua intensitas berbeda (baik 40 menit pada 45,6 ° C (T1) atau 160 menit pada 43,8 ° C (T2)) dibandingkan dengan yang terpapar setengah durasi pada setiap intensitas (yaitu 20 menit pada T1 + 80 menit pada T2), dengan eksperimen menggunakan suhu tinggi baik pertama atau terakhir. Semua kelompok yang stres panas menunjukkan pengurangan pertumbuhan yang signifikan dibandingkan dengan kontrol yang tidak diobati (ANOVA satu arah, p < 0,001 untuk semua kelompok) dan tidak ada perbedaan signifikan yang diamati di antara kelompok perlakuan terlepas dari bagaimana mereka diberi dosis 50% Lt 50 (uji HSD Tukey p > 0,05 untuk semua perbandingan) (Gambar 4 ).
3.4 Dampak berkelanjutan dari stres panas sub-mematikan
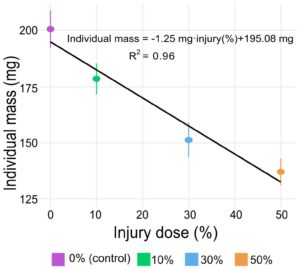
Setelah menetapkan bahwa stres panas subletal mengurangi pertumbuhan dalam cara yang bergantung dosis selama 24 jam setelah stres, kami memeriksa sejauh mana biaya ini bertahan dalam BSFL dalam kondisi yang lebih alami (dan seperti produksi) (yaitu periode pertumbuhan 7 hari setelah stres panas, dengan 100 larva berinteraksi dalam porsi yang sama dari 100 g pakan). Ketika diukur setelah 7 hari, kami mengamati bahwa hampir semua larva bertahan hidup (Kelangsungan hidup >99% di seluruh perlakuan eksperimental). Larva kontrol mencapai massa rata-rata 200 mg ± 7,5 (rata-rata ± SE) setelah 7 hari sementara larva dari kelompok perlakuan stres panas lebih kecil (178 ± 6, 151 ± 6,9 dan 137 ± 5,2 mg (rata-rata ± SE) untuk larva yang menerima 10%, 30% dan 50% dari Lt 50 , masing-masing) (Gambar 5 ). Model linier, dengan dosis stres sebagai faktor, menjelaskan sebagian besar varians dalam berat rata-rata ( R2 = 0,96), yang menunjukkan efek negatif signifikan ( p = 0,023) dari dosis stres panas terhadap massa setelah 7 hari. Dengan demikian, massa larva berkurang 1,25 mg untuk setiap 1% dosis cedera mematikan saat larva diambil sampelnya 7 hari setelah paparan stres. Dihitung sebagai RGR, dibandingkan dengan kelompok kontrol, setiap cedera 1% mengakibatkan pengurangan laju pertumbuhan sebesar 0,14% hari -1 selama periode 7 hari.
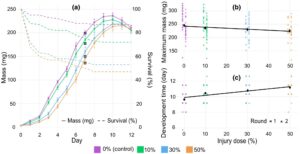
Dalam percobaan akhir, kami mengevaluasi bagaimana stres panas subletal dalam tahap larva awal (instar ke-4 (4 mg)) memengaruhi pertumbuhan selama periode perkembangan hingga larva mencapai tahap prapupa yang tidak makan (Gambar 6 ). Beberapa mortalitas diamati dalam percobaan ini, kemungkinan karena kombinasi stres panas dan penanganan harian. Uji log-rank mengonfirmasi perbedaan signifikan dalam mortalitas antara kelompok perlakuan (Mantel-Cox, p < 0,0001) sehingga mortalitas meningkat dengan intensitas stres panas (Gambar 6a ) (lihat Lampiran S9 ). Yang penting, analisis pertumbuhan hanya mencakup larva yang bertahan hidup, dan penyertaan/pengecualian larva yang sekarat tidak berpengaruh pada pola umum yang diamati (lihat Lampiran S6 ). Seperti yang terlihat pada Gambar 6 , gangguan pertumbuhan yang disebabkan oleh stres panas bertahan pada larva yang dipantau setiap hari selama 12 hari setelah stres panas (Gambar 6a ). Beberapa efek negatif terwujud sebagai pengurangan massa maksimal (Gambar 6b ) dan beberapa dalam perkembangan yang tertunda (Gambar 6c ). Kelompok kontrol mencapai massa maksimum tertinggi sebesar 245,8 ± 5,9 mg (rata-rata ± SE), sementara kelompok yang paling stres (50% dari Lt 50 ) mencapai 224,8 ± 7,2 mg (rata-rata ± SE) (Gambar 6b ). Waktu perkembangan (waktu hingga massa maksimum setelah stres panas) bervariasi dengan tingkat cedera, dengan kelompok perlakuan mencapai massa maksimum pada 9,4, 10,1, 10,4 dan 10,8 hari untuk kontrol, 10%, 30% dan 50% kelompok cedera, masing-masing (Gambar 6c ). Percobaan dilakukan dalam dua putaran percobaan, dan Uji Rasio-Kemungkinan menunjukkan bahwa ‘putaran’ secara signifikan memengaruhi massa maksimum ( p = 0,0013) dan waktu perkembangan ( p = 0,045). Dengan menggabungkan ‘putaran’ sebagai faktor acak, model campuran linear mengungkapkan efek cedera yang signifikan pada kedua sifat tersebut. Oleh karena itu, massa maksimal menurun dengan meningkatnya tingkat cedera ( p < 0,05), dengan pengurangan 0,354 mg per peningkatan 1% dalam dosis mematikan (Gambar 6b ). Waktu perkembangan meningkat dengan tingkat cedera ( p < 0,0001) yang sesuai dengan penundaan 0,0247 hari per peningkatan 1% dalam dosis mematikan (Gambar 6c ). Akhirnya, relatif terhadap kontrol, setiap cedera 1% mengurangi RGR sebesar 0,09% hari -1 , selama periode 10 hari (di mana larva rata-rata telah mencapai massa maksimum).
4 DISKUSI
4.1 Kurva kinerja termal: Batasan untuk kisaran suhu yang diizinkan
Kurva kinerja termal (TPC) sering digunakan untuk menggambarkan bagaimana laju biologis bervariasi dalam berbagai suhu. Luas kurva TPC dapat mencakup suhu yang penuh tekanan ketika mempertimbangkan laju biologis akut seperti laju metabolisme atau laju pergerakan, tetapi di sini kami menggunakan TPC untuk sifat pertumbuhan kronis (jangka panjang), yang hanya dapat tetap positif dalam rentang suhu yang memungkinkan di mana akumulasi bersih stres termal tidak ada (Lihat pembahasan dalam Ørsted et al., 2022 dan Gambar S1 ). Studi sebelumnya mengenai BSF sebagian besar berfokus pada kinerja termal di seluruh siklus perkembangan penuh BSFL. Namun, TPC pertumbuhan larva kami selaras dengan studi BSFL sebelumnya yang melaporkan peningkatan laju pertumbuhan dari 15 °C ke 35 °C, puncak pada 35 °C–37 °C (TPC opt ) dan penurunan dari 35 °C ke 42 °C, dengan pertumbuhan paling efisien dan kelangsungan hidup tinggi terjadi antara 27 °C dan 39 °C (Chia et al., 2018 ; Raimondi et al., 2020 ; Schøn et al., 2024 ). Dalam studi saat ini, kami mengamati penghentian pertumbuhan dan beberapa mortalitas pada ~42 °C, yang karenanya kami identifikasi sebagai proksi untuk suhu kritis ( T C ) di atas mana cedera panas bersih terakumulasi (Ørsted et al., 2022 ; Gambar S1 ). Batas termal kritis untuk BSFL dilaporkan secara ambigu, dengan kelangsungan hidup pada 40°C sebagai rendah (Chia et al., 2018 ; Tomberlin et al., 2009 ) atau tinggi (Purkayastha & Sarkar, 2022 ), sementara Li et al. ( 2024 ) menemukan suhu maksimum kritis (CT maks ) untuk BSFL menjadi ~47°C. Hasil yang berbeda kemungkinan mencerminkan variasi dalam protokol paparan cedera dan metode pengukuran suhu (Ørsted et al., 2022 ). Selain itu, variasi diet dan genetik di antara strain BSFL dapat memengaruhi perkiraan suhu kritis (Chia et al., 2018 ; Purkayastha & Sarkar, 2022 ). Meski begitu, tampaknya paparan kronis pada suhu di atas 42°C akan membuat larva stres.
4.2 Kurva waktu kematian termal: Batasan untuk rentang suhu yang menegangkan
Kisaran suhu yang penuh tekanan mengganggu homeostasis dan menyebabkan akumulasi cedera bersih, yang pada akhirnya menyebabkan mortalitas, seperti yang dijelaskan oleh model waktu kematian termal (TDT) (Jørgensen et al., 2022 ; Ørsted et al., 2022 ; Rezende et al., 2014 ). Dampak paparan panas yang penuh tekanan bergantung pada intensitas (yaitu suhu) dan durasi. Menurut paradigma ini, kegagalan termal mengikuti model yang bergantung pada dosis, yang terjadi setelah dosis cedera yang terbatas (100% dari dosis mematikan) tercapai (Fry, 1946 ; Jørgensen et al., 2021 ). Dalam penelitian saat ini, kami mendefinisikan dosis ini sebagai perawatan stres yang diperlukan untuk mencapai Lt 50 , dan karena suhu yang penuh tekanan yang lebih tinggi mempercepat laju akumulasi cedera, titik akhir kritis tercapai lebih cepat pada suhu yang lebih tinggi (Jørgensen et al., 2021 , 2022 ; Rezende et al., 2014 ). Melalui analisis TDT Lt 50 pada rentang suhu stres di atas T C (42°C–50°C) (Gambar 2 ), kami menemukan korelasi kuat ( R 2 = 0,98) antara log(Lt 50 ) dan suhu pengujian, yang mendukung ekspektasi bahwa laju akumulasi cedera meningkat secara eksponensial seiring suhu (Johnson et al., 2004 ; Jørgensen et al., 2019 , 2022 ; Ørsted et al., 2022 ). Peningkatan eksponensial dalam laju cedera seiring suhu sangat kuat, sehingga peningkatan 1°C akan melipatgandakan laju akumulasi cedera dan karenanya mengurangi waktu toleransi lebih dari setengahnya (Lt 50 ). Dinyatakan sebagai Q 10 (peningkatan faktorial dalam laju biologis untuk kenaikan 10°C), hubungan eksponensial laju akumulasi cedera menghasilkan Q 10 = 1690, yang mencerminkan proses yang sangat sensitif terhadap suhu. Hal ini konsisten dengan median Q 10 (~1500) untuk tingkat kegagalan panas yang baru-baru ini diamati di 112 spesies ektotermik (Jørgensen et al., 2022 ) dan sejalan dengan bukti saat ini dan historis bahwa kegagalan panas ektotermik menunjukkan sensitivitas termal yang ekstrem, dengan nilai Q 10 berkisar dari ~300 hingga ~290.000 (Jørgensen et al., 2019 , 2022 ; Maynard Smith, 1957 ; Ørsted et al., 2022 ). Yang penting, diasumsikan bahwa total dosis stres yang mematikan tetap konstan di berbagai suhu stres dan bahwa pengurangan eksponensial dalam waktu toleransi hanyalah konsekuensi dari peningkatan tingkat cedera yang kuat (Fry, 1946) .; Ørsted dkk., 2022 ).
Sementara model TDT telah memperoleh minat yang besar dalam pemodelan kematian termal (Castañeda et al., 2015 ; Maynard Smith, 1957 ; Truebano et al., 2018 ), kegunaannya dalam mengukur konsekuensi subletal dari stres panas masih kurang dipahami (Ørsted et al., 2024 ; Parratt et al., 2021 ; Walsh et al., 2021 ). Dalam studi saat ini, kami menggunakan dua asumsi model untuk memeriksa efek subletal. Secara khusus, kami berasumsi bahwa kegagalan panas berkembang secara progresif pada suhu yang penuh tekanan, dan kami berasumsi bahwa dosis stres total pada Lt 50 adalah konstan di seluruh rentang suhu yang penuh tekanan (Fry, 1946 ; Ørsted et al., 2022 ). Dengan demikian, dengan memaparkan larva pada sebagian kecil dosis mematikan (Lt50 ) , kita dapat menerapkan ‘beban stres’ yang terkontrol dan submematikan (misalnya 10%, 20%, 30% dari durasi hingga Lt50 pada suhu tertentu). Biaya ‘beban stres’ submematikan spesifik kemudian dievaluasi dalam kondisi yang memungkinkan saat larva dikembalikan dari stres panas ke suhu pertumbuhan optimalnya, dari sana gangguan dalam pertumbuhan dan perkembangan dicatat.
4.3 Mengukur biaya stres panas sub-mematikan
Penelitian sebelumnya yang menggunakan pemodelan TPC telah memodelkan bagaimana campuran suhu optimal dan suboptimal memengaruhi kebugaran rata-rata dalam rentang permisif (di mana kebugaran bervariasi, tetapi tetap positif) (Deutsch et al., 2008 ; Kingsolver et al., 2023 ; Vasseur et al., 2014 ). Di sini kami menggunakan pendekatan yang berbeda untuk memeriksa biaya langsung dari stres panas yang diperoleh di luar batas TPC (yaitu bagaimana periode ‘kebugaran negatif’ atau akumulasi cedera memengaruhi kinerja selanjutnya). Dengan menggabungkan TPC dan TDT—yang mewakili (kembali) pembentukan dan hilangnya homeostasis—kami mengukur biaya stres panas subletal, yang terwujud dalam rentang permisif (pada T opt ) setelah paparan suhu yang penuh tekanan (di atas T C ) (Colinet et al., 2015 ; Ørsted et al., 2022 ). Dalam percobaan pertama kami, kami menguji apakah biaya, diukur sebagai pertumbuhan berkurang satu hari setelah stres, meningkat secara linear dengan dosis cedera (lihat Lampiran S5 untuk hubungan dosis-durasi). Seperti yang dihipotesiskan, peningkatan dosis stres termal menyebabkan pengurangan linear dalam laju pertumbuhan spesifik ketika BSFL dikembalikan ke kondisi pertumbuhan optimal (Gambar 3 ), yang menunjukkan peningkatan proporsional dalam biaya dengan dosis cedera termal. Pengurangan pertumbuhan konsisten untuk dosis cedera tertentu, terlepas dari intensitas yang digunakan (diuji untuk tiga suhu berbeda di atas T C , Gambar 3 ). Selain itu, kami menunjukkan bahwa cedera yang disebabkan oleh panas bergabung secara aditif di berbagai intensitas, terlepas dari urutan paparan (Gambar 4 ). Bersama-sama, hasil ini menunjukkan bahwa jenis cedera yang sama terjadi terlepas dari suhu stres spesifik yang digunakan. Meskipun ini bukan bukti langsung dari proses cedera yang serupa, aditivitas yang diamati dan biaya yang sebanding di seluruh suhu yang penuh tekanan sangat mendukung bahwa akumulasi stres panas di seluruh suhu yang membahayakan berasal dari malfungsi biologis yang serupa, seperti yang sebelumnya diusulkan untuk D. melanogaster (Jørgensen et al., 2021 , 2022 ; Ørsted et al., 2022 ) dan ikan trout berbintik (Fry, 1946 ). Hasil ini menunjukkan bahwa proporsi tertentu dari stres panas yang mematikan menimbulkan biaya tertentu yang kemudian ‘dibayar’ dengan mengalokasikan energi untuk perbaikan dengan mengorbankan pertumbuhan setelah larva dikembalikan ke kondisi yang memungkinkan.
4.4 Biaya stres panas sub-mematikan dalam kondisi seperti produksi
Dalam percobaan kami berikutnya, kami meneliti bagaimana biaya stres terwujud dalam jangka waktu yang lebih lama (7 hari) dengan hipotesis bahwa biaya, seiring waktu, sebagian dan secara bertahap terwujud sebagai perkembangan yang tertunda. Eksplorasi biaya temporal dan energetik dari stres subletal jelas relevan dengan pertanyaan ekologi dan terapan dan BSFL mewakili sistem yang relevan dan praktis untuk mempelajarinya. Dengan demikian, pertumbuhan H. illucens telah memperoleh perhatian yang semakin cepat karena larva ini efisien dalam mengubah limbah organik menjadi biomassa kaya protein dan lemak dalam kondisi kepadatan tinggi (Siddiqui et al., 2024 ; Wang & Shelomi, 2017 ). Untuk mensimulasikan lingkungan pertumbuhan kompetitif untuk BSFL dalam sistem komersial, oleh karena itu, kami meneliti efek subletal dari stres panas selama periode pertumbuhan 7 hari ‘normal’ (Schøn et al., 2024 ). Pada skala waktu ini, kami juga menemukan massa larva menurun secara proporsional dengan meningkatnya fraksi cedera (Gambar 5 ) sehingga kelompok cedera 50% menjadi 32% lebih kecil daripada kelompok kontrol (7 hari setelah paparan stres). Meskipun tingkat kematian rendah dalam percobaan ini, stres panas jelas berdampak pada larva, karena pertumbuhan menurun seiring dengan meningkatnya cedera. Dengan demikian, stres panas pada tahap awal kehidupan mengurangi dan menunda pertumbuhan jangka panjang dengan cara yang dapat diukur dan berbanding lurus dengan dosis stres.
4.5 Biaya ontogenetik dari stres panas sub-mematikan
Dalam rangkaian percobaan terakhir, kami menyelidiki efek berkelanjutan dari paparan panas subletal jangka pendek terhadap pertumbuhan, waktu perkembangan, dan mortalitas pada BSFL yang mengalami perkembangan penuh (hingga prapupa). Ketika dianalisis dari waktu paparan cedera hingga tahap prapupa akhir (berhentinya makan), kami menemukan pengurangan massa sebesar 7,8% pada kelompok cedera 50% dibandingkan dengan kontrol (Gambar 6b ). Oleh karena itu, pengurangan proporsional dalam pertumbuhan lebih kecil ketika diperiksa setelah perkembangan penuh. Namun, kami menemukan bahwa pertumbuhan kompensasi diterjemahkan menjadi waktu perkembangan yang lebih lama. Dengan demikian, waktu perkembangan untuk kelompok yang paling stres (50% dari Lt 50 ) adalah 1,4 hari (11%) lebih lama daripada kelompok kontrol (Gambar 6c ), yang sesuai dengan penundaan 0,0247 hari untuk setiap peningkatan 1% dalam dosis mematikan. Oleh karena itu, hasil kami menunjukkan bahwa, dengan waktu pemulihan yang cukup, stres panas menunda perkembangan sementara keterbatasan dalam massa akhir sebagian pulih. Penundaan perkembangan serupa ditemukan pada nimfa wereng cokelat di mana yang selamat dari uji Lt 50 (menerima 100% Lt 50 ) membutuhkan waktu 23% dan 4% lebih lama daripada kontrol untuk mencapai tahap dewasa untuk jantan dan betina, masing-masing (Piyaphongkul et al., 2012 ). Demikian pula, waktu perkembangan pada kutu daun biji-bijian meningkat setelah stres panas subletal, yang menyebabkan berkurangnya laju pertumbuhan populasi (Jeffs & Leather, 2014 ).
Secara kolektif, data kami menunjukkan bahwa biaya stres panas secara bertahap dibagi menjadi perkembangan yang tertunda dan pertumbuhan yang berkurang dan, yang penting, bahwa besarnya biaya ini secara langsung dan linier terkait dengan dosis stres. Dengan demikian, pendekatan analitis kami memberikan metode yang dapat diukur untuk menilai stres panas subletal. Khususnya, dalam konteks ekologi, kemampuan serangga untuk mengompensasi keterlambatan perkembangan bergantung pada fenologi dan riwayat hidup mereka. Spesies dengan fenologi yang fleksibel dapat pulih dari keterlambatan yang disebabkan oleh stres, sementara spesies dengan pola perkembangan yang disinkronkan lebih rentan terhadap ketidaksesuaian, yang dapat memengaruhi kelangsungan hidup dan reproduksi (Miller-Rushing et al., 2010 ; Parmesan, 2006 ; White & Dillon, 2023 ).
Tingkat kematian yang diamati pada larva individu yang berkembang menjadi pra-pupa (Gambar 6a ) lebih tinggi dari yang diharapkan, mengingat bahwa kami hampir tidak menemukan kematian ketika larva dianalisis setelah 7 hari (Gambar 5 ). Kami berspekulasi bahwa peningkatan kematian ini terkait dengan penanganan larva setiap hari, yang tidak ada selama percobaan 7 hari. Dengan demikian, interaksi aditif atau sinergis antara stres penanganan dan stres panas dapat menjelaskan variasi signifikan dalam kematian di seluruh kelompok cedera, dengan kematian tertinggi pada kelompok yang terluka 50% dan terendah pada kontrol (Gambar 6a ). Yang penting, semua analisis pertumbuhan dan waktu perkembangan dalam percobaan ini hanya mencakup data dari larva yang bertahan hidup hingga tahap prapupa.
Baik kita menganalisis pengurangan pertumbuhan setelah 1 hari, 7 hari atau selama keseluruhan durasi perkembangan BSFL, kita menemukan pengurangan proporsional dalam pertumbuhan relatif terhadap fraksi dosis mematikan (bandingkan Gambar 3 , 5 dan 6b ). Seperti yang dibahas, penekanan fraksional pertumbuhan menjadi kurang jelas dengan meningkatnya waktu pemulihan (lihat Lampiran S10 ). Sementara pertumbuhan awal sangat ditekan, larva masih dapat mencapai massa akhir yang layak dalam kondisi permisif jika mereka diberi waktu dan energi untuk pulih. Mekanisme fisiologis yang mendasari cedera panas dan biaya perbaikan berikutnya pada ektoterm masih kurang dipahami tetapi dapat dikaitkan dengan denaturasi protein, keterbatasan oksigen, hilangnya rangsangan seluler dan disfungsi membran (Cossins & Bowler, 1987 ; González-Tokman et al., 2020 ; Pörtner, 2001 ; Schulte, 2015 ). Dengan demikian, stres panas kemungkinan mengganggu keseimbangan energi, karena memulihkan homeostasis dan memastikan kelangsungan hidup memerlukan realokasi energi dari ciri kebugaran (pertumbuhan, perkembangan, dan reproduksi) menuju proses ketahanan, perbaikan, dan pemeliharaan (Colinet et al., 2015 ; González-Tokman et al., 2020 ). Ekspresi protein syok panas, misalnya, dikaitkan dengan penurunan regulasi fungsi metabolisme basal yang penting, kemungkinan karena proses yang membutuhkan energi untuk degradasi dan pembersihan protein yang salah lipat atau teragregasi yang disebabkan oleh stres panas. (Kingsolver & Woods, 2016 ; Krebs & Feder, 1997 ; Kristensen et al., 2016 ).
5 KESIMPULAN
Ketika suhu global meningkat dan kejadian panas ekstrem meningkat, ektotermik mungkin menghadapi kondisi termal yang mendekati batas termal atas mereka untuk kinerja (Jørgensen et al., 2022 ). Ketika suhu melebihi yang diizinkan untuk penyelesaian siklus hidup, model waktu kematian termal menilai hubungan antara suhu dan waktu toleransi, secara tradisional berfokus pada hasil biner (yaitu kelangsungan hidup vs. mortalitas) (Fry, 1946 ; Rezende et al., 2014 ). Memperkirakan batas termal kritis penting untuk model distribusi spesies yang menilai dampak perubahan iklim (Bennett et al., 2021 ; Sunday et al., 2010 , 2012 ), tetapi pendekatan saat ini mungkin mengabaikan efek subletal, yang bisa dibilang lebih cocok dan memprediksi distribusi spesies global (Ørsted et al., 2024 ; Parratt et al., 2021 ).
Studi saat ini mengukur model TPC dan TDT dalam rentang termal permisif dan stres yang dipisahkan oleh suhu kritis ( T C ) menggunakan BSFL sebagai organisme model. Daripada membahas implikasi langsungnya terhadap populasi BSFL alami, fokus utamanya adalah untuk menunjukkan bahwa stres subletal pada organisme ektotermik dapat menyebabkan konsekuensi fisiologis yang bertahan lama yang dapat diukur dan meningkat secara proporsional terhadap dosis stres setelah hewan dikembalikan ke kondisi permisif. Dengan demikian, kami menyajikan model biaya baru yang mengukur dampak cedera panas subletal pada pertumbuhan serangga. Prasyarat untuk kerangka kerja ini adalah perbedaan antara kurva kinerja termal dan model waktu kematian termal, yang dipisahkan oleh suhu kritis yang didasarkan pada keseimbangan homeostatis (Ørsted et al., 2022 ). Dengan menerapkan parameter TDT, kami mendukung temuan sebelumnya bahwa cedera panas terakumulasi secara aditif di seluruh intensitas suhu hingga dosis mematikan tercapai dan bahwa dosis ini tetap konsisten terlepas dari intensitas stres, dengan suhu yang lebih tinggi hanya memperpendek waktu yang diperlukan untuk mencapainya (Jørgensen et al., 2021 , 2022 ). Cedera panas aditif telah didokumentasikan pada ektotermik (Fry, 1971 ; Jørgensen et al., 2021 ) dan baru-baru ini pada kegagalan termal aktivitas fotosintesis pada tumbuhan (Faber et al., 2024 ). Hasil kami menunjukkan bahwa cedera panas subletal mengurangi pertumbuhan secara proporsional dengan fraksi dosis mematikan yang diterapkan. Dengan waktu pemulihan, efek kompensasi mengurangi pengurangan ini, dengan biaya yang terutama terwujud sebagai penundaan dalam peristiwa riwayat hidup (keterlambatan perkembangan), juga sebanding dengan dosis cedera.
Di sini, kami mengukur stres panas dengan hilangnya pertumbuhan, tetapi percobaan serupa dapat dilakukan dengan sifat-sifat lain, karena stres panas juga berlanjut sebagai biaya metabolik langsung, perubahan perilaku atau penurunan kesuburan (Ma & Ma, 2012 ; Okasha, 1970 ; Ratz et al., 2024 ; Zhu et al., 2017 ). Kami mengusulkan untuk memperluas model stres panas dalam kerangka TDT, karena data kami sangat selaras dengan pemodelan TDT di seluruh suhu yang penuh tekanan. Namun, kehati-hatian diperlukan saat menerapkan data yang diperoleh dari laboratorium pada kondisi alami, di mana dinamikanya lebih kompleks. Suhu mikrohabitat alami kemungkinan berfluktuasi di seluruh T C , dengan kapasitas perbaikan dan akumulasi cedera secara bergantian mendominasi. Model idealnya juga memperhitungkan pergeseran toleransi termal atau resistensi termal dari pengerasan, aklimatisasi, serta mitigasi melalui termoregulasi perilaku, yang semuanya memengaruhi estimasi toleransi (Bowler, 2018 ; González-Tokman et al., 2020 ; Kearney et al., 2009 ; Noer et al., 2022 ). Lebih jauh, penting untuk mengakui bahwa studi ini mengisolasi stres panas, mengecualikan kompleksitas interaksi dengan stresor alami lainnya (misalnya kekeringan, keterbatasan energi atau persaingan intraspesifik), sedangkan penilaian ekologi idealnya memperhitungkan efek gabungannya (Litchman & Thomas, 2023 ).
Temuan yang disajikan di sini relevan untuk pemodelan ekologi umum dari tekanan panas tetapi mungkin juga berlaku langsung dalam kaitannya dengan produksi skala besar larva lalat tentara hitam (BSFL). Dengan demikian, gangguan pertumbuhan dan perkembangan yang tertunda yang terjadi setelah tekanan panas menggarisbawahi perlunya kontrol suhu dalam sistem produksi tersebut untuk mengoptimalkan produktivitas dan konversi energi dari pakan ke larva (Schøn et al., 2024 ). Misalnya, diketahui bahwa aktivitas larva dalam sistem produksi skala besar dapat meningkatkan suhu substrat lebih dari 10°C (Li et al., 2023 ) dan perubahan suhu tersebut dapat melampaui suhu kritis ( T C ) dan akibatnya membatasi produktivitas. Dengan demikian, penelitian kami menyoroti bagaimana paparan subletal terhadap suhu tinggi dalam sistem alami dan komersial dapat mengurangi ukuran hewan, memperpanjang perkembangan dan pada akhirnya menurunkan kebugaran dan/atau pendapatan.