
ABSTRAK
Kami menyajikan, untuk pertama kalinya, perilaku makan dengan cara menghisap dari kecebong Dendropsophus cerradensis (Hylidae, Dendropsophini), bersama dengan deskripsi terperinci tentang morfologi eksternalnya, rongga bukofaringeal, dan sistem muskuloskeletal. Kecebong memperlihatkan tubuh yang cekung, lubang hidung yang terletak di anterior, cakram oral yang dimodifikasi (tertutup sepenuhnya oleh lipatan eksternal), dan ekor yang rendah, menyerupai anggota lain dari kelompok D. microcephalus . Rongga bukofaringeal memiliki ciri-ciri yang mengecil, dengan lubang hidung internal yang terletak pada sudut lancip dan ditutupi oleh papila prenaria, yang hanya ditemukan pada spesies ini. Pola insersi otot secara umum konsisten dengan kecebong Dendropsophini lainnya, kecuali insersi m. levator mandibulae longus profundus pada tulang rawan Meckel. Perilaku makan dicirikan dengan penggunaan tabung oral yang menonjol secara eksklusif selama pemangsaan. Mekanisme ini mungkin terkait dengan otot-otot mandibula dan hyoid yang kuat, serta struktur tengkorak yang dimodifikasi—termasuk elemen suprarostral yang unik, prosesus otot quadrangular, ceratohyal yang kuat, dan keranjang branchial yang berkurang dalam kerangka hyobranchial—yang memungkinkan gerakan hisap yang cepat. Studi ini menyajikan aspek yang sebelumnya tidak diketahui dari tabung oral protractile dan perilaku makan kelompok D. microcephalus , yang memberikan wawasan baru tentang morfologi dan perilaku makan kelompok tersebut.
1 Pendahuluan
Berudu, tahap larva anuran, merupakan komponen ekosistem air tawar yang beragam dan melimpah (misalnya, Inger et al. 1986 ; Ranvestel et al. 2004 ; Whiles et al. 2006 ). Di lingkungan akuatik, berudu mengonsumsi energi sebanyak mungkin untuk berkembang hingga mengalami metamorfosis kompleks (Alford dan Harris 1988 ). Mereka memperoleh nutrisi melalui berbagai strategi, dari sumber makanan yang berbeda, termasuk alga, bahan tanaman, serbuk sari, invertebrata akuatik (telur dan larva), dan bahkan berudu lainnya (Kupferberg 1997 ; Crump 1990 ; 1992; Altig 2007 ; Kloh et al. 2019 ).
Berudu diketahui menunjukkan strategi makan yang beragam. Spesies yang suka mengikis menggunakan struktur berkeratin, seperti selubung rahang dan gigi labial, secara berurutan dan terkoordinasi untuk mengikis serpihan (Venesky et al. 2010 ). Spesies karnivora dapat mencabik mangsanya menggunakan selubung rahang berkeratin sebelum menelannya (Vera-Candioti 2005 ), sementara spesies makrofag menelan seluruh benda dengan cara dihisap (Deban dan Olson 2002 ).
Dendropsophus adalah genus hylid yang paling beragam, dengan 105 spesies yang dikenali yang ditugaskan ke delapan kelompok spesies (Faivovich et al. 2005 ; Orrico et al. 2021 ; Whitcher et al. 2025 ; Frost 2025 ). Kelompok Dendropsophus microcephalus saat ini terdiri dari 35 spesies (Orrico et al. 2021 ; Nakamura et al. 2025 ). Kelompok ini pertama kali ditetapkan untuk mengakomodasi spesies yang berudunya memiliki ekor xiphicercal dan cakram oral terminal, tidak memiliki gigi dan papila labial (Duellman 1970 ). Berudu diklasifikasikan dalam serikat ekomorfologi makrofag, yang dicirikan oleh LTRF 0/0 atau 0/1; adanya selubung rahang; adanya cakram oral terminal kecil dengan sedikit atau tidak ada papila marginal (Altig dan Johnston 1989 ).
Cakram oral termodifikasi dari kelompok Dendropsophus microcephalus telah dideskripsikan dengan berbagai cara dalam literatur, tetapi secara konsisten dicirikan oleh tidak adanya gigi labial dan/atau papila marginal. Awalnya, Bokermann ( 1963 ) mendeskripsikannya sebagai mangkuk penghisap berbentuk U pada D. nanus . Kemudian, Lavilla ( 1990 ) mengidentifikasinya sebagai tabung penghisap berbentuk U yang dapat ditarik. Deskripsi lain tidak menyebutkan struktur tersebut atau mendeskripsikannya hanya sebagai cakram oral ventral kecil, seperti pada D. microcephalus (Cruz dan Dias 1990 ) dan D. studerae (Carvalho-e-Silva et al. 2003 ). Vera-Candioti et al. ( 2004 ) menyebutnya sebagai tabung penghisap. Deskripsi terbaru D. branneri menyoroti struktur berbentuk U (De Abreu et al. 2015 ). Kaplan ( 2017 ), berdasarkan potongan histologis, memberikan deskripsi rinci tentang struktur pada D. microcephalus , D. rhodopeplus , dan D. mathiassoni .
Penonjolan diskus oral pada kelompok D. microcephalus telah banyak dibahas dalam literatur. Lavilla ( 1990 ) adalah orang pertama yang mengenali retraksi diskus oral. Vera-Candioti et al. ( 2004 ) membahas bagaimana sistem muskuloskeletal dapat berfungsi dalam pemberian makan dengan cara menghisap pada D. nanus dan kemudian menyarankan fungsi penonjolan (Vera-Candioti 2007 ). Meskipun Kaplan ( 2017 ) menolak gagasan tentang ekstensi dan retraksi diskus oral karena labium bawah yang turgid, ia mengakui kemungkinan proyeksi tuba jika lipatan eksternal dapat ditarik.
Analisis sebelumnya tentang isi usus mendukung diet makrofag pada berudu kelompok D. microcephalus . Wassersug dan Rosenberg ( 1979 ) menemukan bahan tumbuhan yang besar dan berlimpah dalam usus D. phlebodes dan D. microcephalus . Selanjutnya, oligochaetes utuh dideskripsikan sebagai makanan utama, diikuti oleh alga yang lebih kecil, pada D. microcephalus dan D. nanus (Vera-Candioti et al. 2004 ; Vera-Candioti 2007 ). Sementara spesies ini mungkin menunjukkan preferensi untuk oligochaetes, data dari Wassersug dan Rosenberg ( 1979 ) juga menunjukkan fleksibilitas dalam diet mereka.
Morfologi internal—termasuk volume bukal yang besar, filter insang yang berkurang, dan tidak adanya epitel sekretori pada Dendropsophus microcephalus (Wassersug dan Hoff 1979 )—juga mendukung dimasukkannya kecebong dari kelompok Dendropsophus microcephalus dalam serikat ekomorfologi makrofag. Selain itu, sistem muskuloskeletal diketahui untuk Dendropsophus nanus dan D. microcephalus (Vera-Candioti 2007 ), dan rongga bukofaringeal telah dideskripsikan untuk D. nanus , D. microcephalus , dan D. phlebodes (Wassersug 1980 ; Vera-Candioti 2007 ). Otot tubuhnya kuat (yakni otot hyoid dan mandibular), tengkorak larva sempit, keranjang branchial dan fitur buccopharyngeal sangat berkurang, dan ceratohyal kuat (Wassersug 1980 ; Vera-Candioti 2007 ), semua fitur tersebut terkait dengan konsumsi barang-barang besar.
Dendropsophus cerradensis (Napoli dan Caramaschi 1998 ) adalah katak pohon kecil dari kelompok Dendropsophus microcephalus (Faivovich et al. 2005 ; Orrico et al. 2021 ; Whitcher et al. 2025 ). Meskipun ada beberapa informasi terfragmentasi yang tersedia tentang kecebongnya, deskripsi formal masih belum ada (lihat Alves-Ferreira et al. 2021 ), dan fitur morfologi internalnya tidak diketahui. Di sini, kami menyajikan analisis morfologi eksternal, rongga bukafaring, dan sistem muskuloskeletal, dan menggambarkan perilaku makan hisap kecebong Dendropsophus cerradensis . Selain itu, kami membahas perilaku makan spesies ini, membandingkannya dengan kecebong makrofag lainnya untuk memberikan wawasan tentang mekanisme makan hisap dalam kelompok Dendropsophus microcephalus .
2 Bahan dan Metode
2.1 Pengambilan sampel
Kami mengumpulkan berudu dan dewasa di Refúgio Estadual da Vida Silvestre Libélulas da Serra de São José (21°06′52.7″S; 44°12′13.7″W), di kotamadya Tiradentes, dan berudu di Parque Nacional da Serra da Canastra (20°09′56.4″S 46°41′15.7″W), di kotamadya São Roque de Minas, keduanya terletak di negara bagian Minas Gerais, Brasil tenggara, di area terbuka dalam domain Cerrado (Gambar 1 ; informasi tambahan di Informasi Pendukung). Kami menidurkan berudu dan dewasa dengan lidokain 5% dan memfiksasinya dalam formalin 10%. Kecebong diawetkan dalam formalin, sementara berudu dewasa disimpan dalam etanol 70%. Selain itu, kami mengawetkan satu kecebong dari setiap lot dalam etanol absolut untuk identifikasi molekuler. Kami menyimpan lot dan spesimen di Koleksi Kecebong dan Dewasa (UFMG-GIR dan UFMG-AMP) dari Centro de Coleções Taxonômicas dari Universidade Federal de University of Minas Gerais (CCT-UFMG) dan Koleksi Amphibia-kecebong dari Universidade Estadual Júlio de Mesquita, kampus São José do Rio Preto (DZSJRP-Amphibia-kecebong; Tabel S1 ). Identifikasi taksonomi dari kecebong dan dewasa dikonfirmasi melalui penanda molekuler mitokondria (Informasi Pendukung).

2.2 Deskripsi Kecebong
Deskripsi morfologi eksternal, pengukuran dan proporsi didasarkan pada sembilan kecebong pada tahap 28–32 (sensu Gosner 1960 ; Tabel S1 ). Kami mengikuti Altig dan McDiarmid ( 1999 ) dan Altig ( 2007 ) untuk terminologi morfologi. Kami mengambil 24 pengukuran hingga 0,1 mm terdekat dari foto mengikuti Altig dan McDiarmid ( 1999 ), Grosjean ( 2005 ), Lavilla dan Scrocchi ( 1986 ), Pinheiro et al. ( 2012 ), dan Lins et al. ( 2018 ) (Informasi Pendukung). Kami mengikuti Pezzuti et al. ( 2021 ) untuk standardisasi karakter dan status karakter dan Schlosser ( 2002a , 2002b ) untuk terminologi garis lateral. Kami memperoleh foto-foto standar dari pandangan lateral, dorsal, dan ventral spesimen menggunakan platform yang dapat disesuaikan untuk menopang kecebong yang terendam dalam air (Schacht dan McBrayer 2009 ). Mengenai analisis morfometrik, kami menggunakan TpsUtil 1.78 (Rohlf 2019 ) untuk kompilasi gambar, TpsDig 2.31 (Rohlf 2017 ) untuk pengukuran, dan ImageJ Versi 1.50b (Schneider et al. 2012 ) untuk pengukuran sudut.
Lima spesimen kecebong digunakan untuk analisis histologis. Hewan-hewan tersebut difiksasi dalam larutan formalin berbuffer, didehidrasi dalam rangkaian etanol bertahap, diafanisasi dalam xylol, ditanamkan dalam parafin Histosec (Merck) dalam Leica Enclosed System (TP1020, Leica Biosystems), dan kemudian dipotong dengan ketebalan 6–8 mikrometer menggunakan mikrotom putar otomatis. Untuk dokumentasi fotomikrografi, digunakan irisan setebal 6 mikrometer. Kecebong ditanamkan untuk memperoleh irisan sagital hewan. Setelah deparafinisasi dan rehidrasi, irisan histologis diwarnai menggunakan teknik hematoxylin-eosin. Slide difoto menggunakan fotomikroskop cahaya (Olympus BX60). Gambar didigitalisasi menggunakan perangkat lunak Image-Pro Plus 6.1 (Media Cybernetics, Silver Spring, MD). Gambar digital diproses dengan Adobe Photoshop dan Adobe Illustrator. Kami mengikuti Altig dan McDiarmid ( 1999 ), Altig ( 2007 ), dan Kaplan ( 2017 ) untuk terminologi morfologi.
Kami membedah dua individu (stadium 32 dan 37, Tabel S1 ) menurut Wassersug ( 1976a ) untuk mengekspos rongga bukafaring dan menerapkan protokol Dias dan Anganoy-Criollo ( 2024 ) untuk mikroskop elektron pemindaian (SEM). Kami mengikuti Wassersug ( 1976a ) untuk terminologi deskriptif. Untuk studi kranium larva dan sistem muskuloskeletal, kami memproses dua individu (stadium 31–32) menurut protokol Wassersug ( 1976b ) untuk pembersihan dan pewarnaan ganda; kami menghentikan prosedur setelah langkah Alcian Blue dan membedahnya secara manual untuk studi otot larva. Setelah pengamatan dan ilustrasi, kami menyelesaikan protokol pembersihan. Kami mengikuti Vera-Candioti et al. ( 2024 ) untuk terminologi kranium dan otot larva dengan nama dalam bahasa Inggris, jika tersedia. Untuk prosedur ini kami mengambil foto dengan mikroskop stereo Leica M205A.
2.3 Perilaku Makan dan Pergerakan Diskus Oral
Di laboratorium, sekitar 2 jam setelah pengumpulan, kami menempatkan kecebong dan vegetasi yang muncul di akuarium yang sama dengan air bersih, dan memfilmkannya dengan kamera Canon D60, pada kecepatan 60 bingkai (masing-masing 0,016 d) per detik selama satu setengah jam. Kami melakukan pengamatan di mana oligochaetes, yang diperoleh di lokasi yang sama dengan kecebong, terus menerus ditawarkan. Kami mendasarkan deskripsi perilaku makan dan mekanika cakram oral pada peristiwa hisapan tunggal yang direkam pada pukul 00:30 pagi (Informasi Pendukung, Video S1 ). Setelah merekam, kami melakukan eutanasia pada kecebong seperti yang dijelaskan di bagian pengambilan sampel.
3 Hasil
3.1 Identifikasi Kecebong
Urutan kecebong dan dewasa tidak menunjukkan perbedaan nukleotida, yang mengonfirmasikan asosiasi semaforontik mereka. Urutan kami menunjukkan 99,7% identitas dengan urutan referensi Dendropsophus cerradensis ( sensu Nakamura et al. 2025 ) yang tersedia di Genbank (nomor akses: MT503752–53) (detail dalam Informasi Pendukung dan Tabel S2 ).
3.2 Morfologi Eksternal
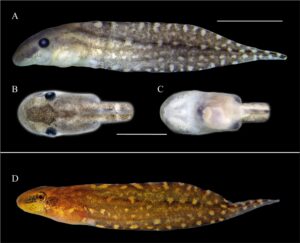
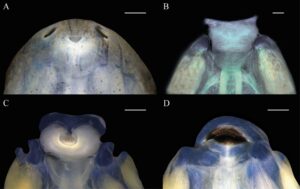
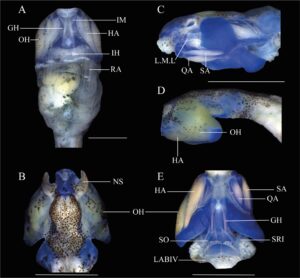
Badan tertekan (Gambar 2 ) (BH/BW 0,76–0,88), elips memanjang dengan penyempitan medial pada pandangan dorsal, segitiga tertekan pada pandangan lateral; tinggi badan sekitar 50% dari panjang badan (BH/BL 0,43–0,50). Moncong membulat pada pandangan dorsal (BWN/BWE 0,33–0,39) dan lateral. Mata diposisikan secara lateral (IOD/BWE 0,85–0,88) diarahkan secara dorsolateral, sekitar 25% dari lebar badan (ED/BWE 0,22–0,26). Lubang hidung elips, diposisikan dan diarahkan secara anterior, mewakili sekitar 5% dari panjang badan (ND/BL 0,04), terlihat secara lateral dan ventral, terletak kira-kira pada diskus oral, yang sesuai dengan sekitar 70% dari lebar diskus oral (ND/ODW 0,50–0,80). Sel bersilia di sekitar lubang hidung, terkonsentrasi di tepi luar (Gambar 3 ). Spirakel sinistral, terlihat dalam pandangan lateral (SDV/BH 0,37–0,64), berukuran kecil (SL/BL 0,09–0,13), diarahkan ke posterior; dinding bagian dalam menempel pada tubuh dan sedikit lebih panjang dari dinding luarnya; lubang terletak pada sepertiga kedua dari panjang tubuh. Tabung intestinal melingkar, titik putar terkilir dari bagian tengah daerah abdomen. Tabung ventilasi dekstral, diarahkan ke posterior, pendek, menempel dan diposisikan di atas tepi sirip ventral; dinding ventral lebih panjang dari dinding dorsal. Diskus oral termodifikasi menjadi tabung oral (Gambar 4 ), anteroventral (ODP 36°), sekitar 11% dari lebar tubuh (ODW/BW 0,07–0,14), papila marginal dan submarginal tidak ada; LTRF 0/0; selubung rahang lebar, bergerigi halus; selubung rahang atas berbentuk lengkung, selubung rahang bawah berbentuk U; selubung tidak tampak saat diskus oral ditarik. Lubang oral kecil, ditutupi oleh lipatan eksternal atas dan bawah yang berkembang dengan baik, yang merupakan kelanjutan dari kulit rostral dan gular (Gambar 3 , 5 dan 5 ). Lipatan eksternal bawah dengan sel bersilia (Gambar 3 ). Lipatan eksternal tersusun dari jaringan ikat halus dan longgar. Labia atas dan bawah ada. Labium atas pendek dan tipis, sedangkan labium bawah didukung oleh jaringan ikat yang lebih panjang dan lebih tebal. Sebuah tonjolan atas terletak di antara labium atas dan selubung rahang atas. Permukaan distal labium bawah cekung dan labium atas cembung. Ekor rendah (MTH/TAL 0,25–0,30), lebih tinggi dari badan (MTH/BH 1,32–1,57), tinggi otot ekor sekitar 30% dari tinggi badan (TMH/BH 0,30–0,34), mencapai ujung ekor; ujung ekor berada di flagel yang jelas. Sirip punggung dan sirip perut memiliki tinggi sedang (DFH/TAL 0,09–0,16 0,12 ± 0,02, VFH/TAL 0,06–011 0,08 ± 0,01). Sirip punggung lebih tinggi dari sirip perut (DFH/VFH 1,30–1,73), berawal di sepertiga bagian belakang badan. Sirip perut kurang melengkung dibandingkan sirip punggung, berawal di tingkat tabung ventilasi. Sistem garis lateral berbeda hanya pada garis supraorbital, dimulai dari posteromedial ke mata dan bertemu ke arah dorsal ke arah moncong, dengan konsentrasi lima baris yang masing-masing berjumlah sekitar 26 jahitan di atas cakram oral dan di antara lubang hidung
3.3 Pengukuran
Tabel 1 .
| Pengukuran | Panggung | Berarti | SD | menit | Maksimal | |||
|---|---|---|---|---|---|---|---|---|
| 28 (n=1) | 30 (n=3) | 31 (n=3) | 32 (n=2) | |||||
| Bahasa Inggris | 16.5 | 20,8 ± 1,0 | 19,1 ± 1,4 | 22,3 ± 2,0 | 19.7 | 2.5 | 16.5 | 22.3 |
| Bahasa Inggris | 4.8 | 6,1 ± 0,7 | 5,9 ± 0,4 | 6,8 ± 0,3 | 5.9 | 0.8 | 4.8 | 6.8 |
| BICARA | 11.6 | 14,7 ± 0,3 | 13,2 ± 1,1 | 15,5 ± 1,7 | 13.8 | 1.7 | 11.6 | 15.5 |
| Bahasa Inggris | 3.1 | 4 ± 0,3 | 3,7 ± 0,2 | 4,3 ± 0,0 | 3.8 | 0.5 | 3.1 | 4.3 |
| DFH | 1.2 | 1,6 ± 0,2 | 1,4 ± 0,2 | 1,8 ± 0,0 | 1.5 | 0.2 | 1.2 | 1.8 |
| VFH | 0.8 | 1,2 ± 0,2 | 0,9 ± 0,2 | 1,3 ± 0,0 | 1.0 | 0.2 | 0.8 | 1.3 |
| TMH | 1.6 | 2 ± 0,1 | 1,9 ± 0,2 | 2,1 ± 0,1 | 1.9 | 0.2 | 1.6 | 2.1 |
| BH | 2.2 | 2,7 ± 0,3 | 2,7 ± 0,0 | 3,1 ± 0,2 | 2.7 | 0.3 | 2.2 | 3.1 |
| Bahasa Inggris | 0.5 | 0,7 ± 0,1 | 0,7 ± 0,1 | 0,7 ± 0,0 | 0.6 | 0.1 | 0.5 | 0.7 |
| Bahasa Indonesia: SVD | 1.4 | 1,5 ± 0,4 | 1,5 ± 0,1 | 1,5 ± 0,4 | 1.5 | 0.1 | 1.4 | 1.5 |
| Bahasa Inggris SED | 3.6 | 4,5 ± 0,5 | 4,6 ± 0,2 | 4,6 ± 0,2 | 4.3 | 0.5 | 3.6 | 4.6 |
| EDI | 0.7 | 0,8 ± 0,1 | 0,8 ± 0,0 | 1 ± 0,1 | 0.8 | 0.1 | 0.7 | 1.0 |
| BW | 2.9 | 3,4 ± 0,3 | 3,2 ± 0,0 | 3,7 ± 0,5 | 3.3 | 0.3 | 2.9 | 3.7 |
| BWN | 1.1 | 1,2 ± 0,0 | 1,2 ± 0,1 | 1,4 ± 0,0 | 1.2 | 0.1 | 1.1 | 1.4 |
| Bahasa Inggris BWE | 2.9 | 3,5 ± 0,2 | 3,4 ± 0,2 | 3,9 ± 0,2 | 3.4 | 0.4 | 2.9 | 3.9 |
| Saat ini | 1.1 | 1,5 ± 0,1 | 1,4 ± 0,1 | 1,7 ± 0,2 | 1.4 | 0.2 | 1.1 | 1.7 |
| ESD | 2.4 | 3,1 ± 0,3 | 3 ± 0,1 | 3,1 ± 0,1 | 2.9 | 0.3 | 2.4 | 3.1 |
| Bahasa Indonesia: NSD | 0.5 | 0,6 ± 0,0 | 0,6 ± 0,0 | 0,6 ± 0,1 | 0.6 | 0.0 | 0.5 | 0.6 |
| ND | 0.3 | 0,3 ± 0,0 | 0,3 ± 0,0 | 0,3 ± 0,0 | 0.3 | 0.0 | 0.3 | 0.3 |
| IND | 0.9 | 1 ± 0,1 | 1 ± 0,1 | 1,1 ± 0,0 | 1.0 | 0.1 | 0.9 | 1.1 |
| IOD | 2.5 | 3,1 ± 0,1 | 3 ± 0,2 | 3,4 ± 0,2 | 3.0 | 0.4 | 2.5 | 3.4 |
| ODW | 0.4 | 0,4 ± 0,0 | 0,4 ± 0,1 | 0,4 ± 0,0 | 0.4 | 0.0 | 0.4 | 0.4 |
| DFiA | 175,1° | 171 derajat ± 7,7 derajat | 167,2 derajat ± 6,0 derajat | 161,3° ± 7,5° | 168,7° | 5,9° | 161,3° | 175,1° |
| ODP | 40,1° | 40,5 derajat ± 6,3 derajat | 28,5° ± 3,2° | 38,3 derajat ± 0,9 derajat | 36,8° | 5,7° | 28,5° | 40,5° |
Catatan: Panjang total (TL), panjang badan (BL), panjang ekor (TAL), tinggi ekor maksimum (MTH), tinggi sirip punggung (DFH), tinggi sirip perut (VFH), tinggi badan (BH), panjang spirakel (SL), jarak spirakel-venter (SVD), jarak moncong-spirakel (SED), diameter mata (ED), lebar badan (BW), lebar badan setinggi lubang hidung (BWN), lebar badan setinggi mata (BWE), lebar otot ekor (TMW), jarak mata-moncong (ESD), jarak lubang hidung-moncong (NSD), diameter narial (ND), jarak internari (IND), jarak interorbital (IOD), lebar cakram oral (ODW), sudut penyisipan sirip punggung (DFIA), posisi cakram oral (ODP). Semua ukuran dalam milimeter (mm) kecuali untuk DFiA dan ODP yang dalam derajat, nilai dinyatakan sebagai mean ± standar deviasi (SD) (rentang) untuk setiap tahap dan untuk pengukuran.
3.4 Warna
Saat hidup, tubuh bercorak bintik-bintik emas dan putih. Jika dilihat dari punggung, bintik cokelat di bagian tengah setinggi otak. Secara lateral, garis memanjang berwarna cokelat memanjang dari mata ke moncong, memudar di bagian depan; iris dengan garis memanjang oranye-cokelat dengan bercak emas di bagian atas dan bawah; permukaan ventral, di bawah garis mata ke perut, berwarna krem dengan bintik-bintik emas di area keranjang insang; perut berwarna perak. Ekor, termasuk otot dan sirip, bercorak homogen dengan bintik-bintik cokelat tua dan emas yang membulat. Sirip tembus cahaya di ujung ekor. Saat diawetkan, warnanya memudar, tetapi bintik melanofor tetap sama.
3.5 Rongga Bukofaringeal
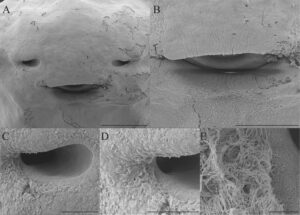
Dasar bukal berbentuk segitiga dengan sepasang papila infralabial bulat yang berkembang baik di bagian tengah dengan tepi yang tidak rata (Gambar 6 ). Anlage lidah berkembang baik. Papila lingual tidak ada. Arena dasar bukal tidak jelas, dengan beberapa pustula di dekat bagian lateral dasar bukal. Kantong bukal tidak terlihat. Zona kelenjar tidak ada. Velum ventral halus, tanpa lubang sekretori dan tidak didukung oleh spikula.
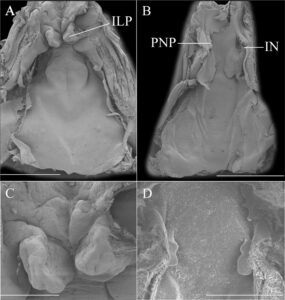
Atap bukal berbentuk segitiga. Lubang hidung bagian dalam berposisi 34 derajat terhadap sumbu longitudinal, sebagian ditutupi papila prenaria (Gambar 6 ). Tidak ada struktur lain yang ada.
3.6 Otot
Tiga puluh satu otot kranial diidentifikasi pada berudu D. cerradensis (Gambar 7 ; Tabel 2 ). Sebagian besar otot mengikuti pola insersi umum (Vera-Candioti 2007 ; Dias et al. 2019 ; Dias et al. 2023 ), dengan beberapa pengecualian yang telah dijelaskan untuk spesies terkait lainnya (yaitu, levator mandibulae lateralis berinsersi pada kantung hidung; kelompok mandibula dan hyoid kuat, terutama m. levator mandibulae longus superfisialis dan profundus, mlm internus, m. orbitohyoideus, dan m. hyoangularis; m. subarcualis rectus II–IV terputus dengan dua slip, dari ceratobranchial I ke prosesus brankial III dan dari prosesus brankial III ke ceratobranchial IV). Konfigurasi m. levator mandibulae longus profundus yang menyisipkan pada tulang rawan Meckel dijelaskan pertama kalinya pada suku tersebut.
| Otot | Asal | Insersi | Komentar |
|---|---|---|---|
| Kelompok mandibula, n. trigeminus (cn V) dipersarafi | |||
| Levator mandibula longus superfisialis | Batas luar dan posterior dari bar subokular dan bagian dari asenden | Daerah dorsomedial tulang rawan Meckel | Pemasangan melalui tendon panjang |
| Levator mandibula longus profundus | Batas medial bar subokular | Batas medial internal tulang rawan Meckel | Penyisipan melalui serat pakan |
| Levator mandibula eksternus superfisialis | Permukaan medial, inferior dari prosesus muskularis | Permukaan medioventral suprarostral | |
| Levator mandibula eksternus profundus | Permukaan medial, inferior dari prosesus muskularis | Margin ventral dari tepi suprarostral | |
| Levator mandibula artikularis | Bagian bawah permukaan medial prosesus muskularis | Permukaan dorsal tepi lateral tulang rawan Meckel | |
| Otot levator mandibula lateralis | Proses artikular palatoquadrate | Kantung hidung | |
| Tulang rahang bawah | Daerah medial tulang rawan Meckel | Aponeurosis median | |
| Mandibula bilier | Daerah ventromedial tulang rawan Meckel | Permukaan bawah, medial dan lateral tabung oral | |
| Otot levator mandibula internus | Permukaan ventral dari prosesus asendens | Tepi bagian dalam tulang rawan Meckel | Pemasangan melalui tendon yang panjang dan sangat tipis |
| Grup hyoid, n. facialis (cn VII) | |||
| Hyoangularis | Permukaan anterior tepi seratohyal | Proses retroartikular tulang rawan Meckel | |
| Kuadrat sudut | Permukaan ventral palatoquadrate | ||
| Suspensorium angularis | Batas ventral prosesus muskularis | Proses retroartikular tulang rawan Meckel | |
| Orbitohyoideus | Batas anterior dan dorsal dari prosesus muskularis | Tepi lateral seratohia | |
| Suspensoriohyoideus | Batas posterior desenden dari prosesus muskularis | Permukaan posterior prosesus lateralis seratohyal | |
| Tulang rawan interhioid | Permukaan ventral ceratohyal, dekat tepi lateral | Aponeurosis median | |
| Kelompok brankial, n. glossopharyngeus (cn IX) dan vagus (cn X) | |||
| Otot levator arcuum branchialium I | Batas lateral bar subokular | Ceratobranchial I | |
| Levator arcuum brankialium II | Proses otik larva | Ceratobranchial I dan Ceratobranchial II | |
| Levator arcuum brankialium III | Bagian lateral kapsul telinga | Ceratobranchial III | |
| Levator arcuum brankialium IV | Bagian posterolateral kapsul otik | Ceratobranchial IV dan lempeng hipobranchial | |
| Timpanofaring | Bagian lateral kapsul telinga | Jaringan ikat anterior terhadap glotis | |
| Dilator laring | Permukaan posterolateral kapsul otik | Tulang rawan aritenoid | |
| Konstriktor branchialis II | Proses insang II | Komisura terminal I | |
| Konstriktor branchialis III | Proses insang II | Komisura terminal II | |
| Konstriktor branchialis IV | Komisura terminal II dan bagian distal ceratobranchial III | Serat-seratnya berkesinambungan dengan serat-serat bagian anterior m. subarcualis rectus II–IV | |
| Otot rektus subarcualis I | Dua slip: bagian lateral dari proses posterior ceratohyal – proksimal, bagian lateral ceratobranchial I (slip dorsal) dan proses branchial III (slip ventral) | ||
| Otot rektus subarkualis II–IV | Dua slip: bagian anterolateral dari ceratobranchial I – proses branchial III (slip anterior) dan proses branchial III – bagian ventromedial dari ceratobranchial IV (slip posterior) | ||
| Otot subarkualis miring II | Tepi posterior basibranchial | Prosesus insang II dan III | |
| diafragma brankialis | Peritonium (rongga perut) | Tepi distal ceratobranchial III | |
| Kelompok tulang belakang, persarafan saraf tulang belakang | |||
| Geniohyoideus | Permukaan posterior, ventral infrarostral | Lempeng hipobranchial, dekat tepi anterior | |
| Otot rektus abdominis | Sabuk panggul | Insersi anterior di ujung anterior lipatan eksternal | |
| Rektus serviks | Peritonium (rongga perut) | Proses insang III | |
Tabel 2 .
3.7 Tengkorak Larva
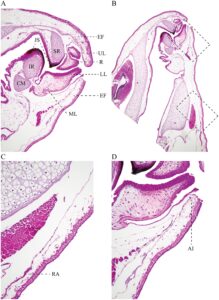
Tidak ada osifikasi kranial pada stadium yang diperiksa (Gambar 8 ). Kranium larva lebih panjang daripada lebar. Suprarostral sebagai satu bagian tulang rawan, hampir lurus; batas proksimal menurun ke arah garis tengah. Bagian tertinggi pada tingkat proses posterior. Tulang rawan suprarostral berartikulasi secara sindesmotik dengan tanduk trabekular. Tanduk trabekular pendek, runcing, melengkung ke bawah di anterior. Dalam pandangan dorsal, kedua tanduk hampir sejajar. Margin medial mengarah ke dalam. Ligamentum quadratoethmoid menghubungkan prosesus lateralis trabekula dan proses quadrato ethmoidal. Dasar kranial sedikit terkondronifikasi di anterior. Fenestra basicranial tidak tersumbat; planum intertrabecular tidak berkembang di daerah medial; dasar kranial ditusuk oleh foramen karotis. Foramen kraniopalatina tidak terlihat. Tulang rawan orbital tinggi, dengan dua lekukan, mengasumsikan batas berbentuk W. Foramen prootik terlihat. Tengkorak larva terbuka ke arah dorsal; fenestra frontoparietal dibatasi oleh taeniae tecti di bagian marginal dan di bagian posterior oleh tectum synoticum. Kapsul telinga berbentuk segi empat dan kuat; kedua kapsul dijembatani oleh tectum synoticum.
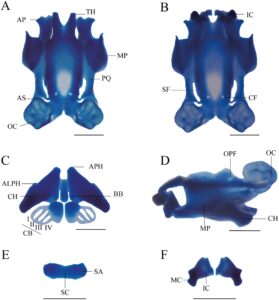
Palatoquadrate panjang dan kuat. Prosesus artikular kuat dengan tepi luar anterior menjorok ke atas. Prosesus pseudopterygoideus tidak ada. Prosesus muskularis lebar, berbentuk segi empat dengan prosesus triangular anterior di tepi anterior. Palatoquadrate menempel pada tempurung otak melalui komisura quadratocranial, prosesus ascendens, dan prosesus otic larval. Komisura quadratocranial lebar. Batang subokular tipis dan melengkung. Prosesus ascendens menempel pada dasar tengkorak. Tulang rawan Meckel lebar, berbentuk sigmoid, berorientasi melintang. Tulang rawan infrarostral pendek, independen, bergabung secara sindesmotik di bagian medial.
Ceratohyal berbentuk segitiga, melengkung relatif terhadap sumbu sagital dan kuat; prosesus anterior, prosesus lateral, kondilus artikular membulat dan kuat, dan prosesus anterolateral kecil dan hampir tidak terlihat. Tepi ceratohyal medial sejajar. Basihyal tidak ada. Basibranchial panjang dan tidak memiliki prosesus urobranchial. Pars reuniens kurang berkondronifikasi. Keranjang branchial mengecil, kurang berkondronifikasi (sulit untuk divisualisasikan dalam spesimen yang telah disiapkan) dengan empat ceratobranchial. Pelat hyobranchial berkembang dengan baik, berbentuk segi empat, diartikulasikan dengan celah medial. Ceratobranchial tidak memiliki proyeksi dan spikula. Ceratobranchial dihubungkan oleh komisura terminal.
3.8 Perilaku Makan dan Pergerakan Diskus Oral
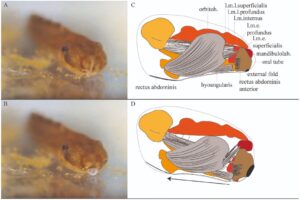
Kecebong sebagian besar waktu tetap tenang di dasar akuarium. Mereka tidak secara aktif mengejar cacing di kolom air, dan untuk beberapa saat, mereka membengkokkan tubuh ke bawah ke arah mangsa. Kecebong bergerak di sepanjang dasar akuarium dengan membengkokkan tubuh, tanpa menggerakkan ekor. Peristiwa predasi yang ditangkap dalam pandangan frontal memperhatikan kecebong pada tahap 36 memangsa cacing yang bergerak di depan moncongnya (Video dalam Informasi Pendukung, Gambar 9 ). Awalnya, gerakan depresi tubuh diamati sepanjang garis moncong longitudinal. Kemudian, moncong mulai tertekan. Akhirnya, tabung mulut sepenuhnya menonjol, dan cacing telah tersedot masuk. Ketiga gerakan awal ini terjadi dalam 0,048 detik, masing-masing berlangsung 0,016 detik. Dalam frame berikutnya, kecebong bergerak maju dan kemudian menyedot cacing melalui serangkaian gerakan terkoordinasi yang melibatkan tabung mulut dan rahang bawah. Setelah 0,13 detik, gerakan terakhir adalah tindakan penghisapan lengkap menggunakan tabung oral. Setelah labia ditarik sepenuhnya, rahang bawah terus bergerak. Seluruh peristiwa pemangsaan, dari gerakan pertama hingga akhir, berlangsung selama 2,56 detik. Karena kamera merekam pada 60 bingkai per detik, setiap bingkai berlangsung selama 0,016 detik, gerakan sebenarnya mungkin terjadi lebih cepat daripada yang diamati.
4 Diskusi
Morfologi larva tampaknya dilestarikan dalam kelompok Dendropsophus microcephalus . Misalnya, semua kecebong yang dikarakterisasi yaitu D. bipunctatus , D. branneri , D. leali , D. mathiassoni , D. meridianus , D. microcephalus , D. nanus , D. phlebodes , D. rhodopeplus, D. robertmertensi , D. rubicundulus , D. sanborni , D. sartori , dan D. studerae memiliki tubuh yang memanjang dan tertekan, mata lateral, cakram oral tanpa papila dan deretan gigi labial yang dimodifikasi dalam tabung oral, lubang hidung terletak di ujung moncong dekat cakram oral, ekor dan sirip rendah, ujung flagel yang jelas, dan garis coklat anteroventral yang membentang dari lubang hidung hingga ujung tubuh (Duellman dan Fouquette 1968 ; Duellman 1972 ; Bokermann 1963 ; Altig 1987 ; Hero 1990 ; Cruz dan Dias 1990 ; Pugliese dkk. 2001 ; Carvalho-e-Silva dkk. 2003 ; Lynch 2006 ; Lynch dan Suárez-Mayorga 2011 ; Schulze dkk. 2015 ; De Abreu dkk. 2015 ).
Cakram oral yang dimodifikasi menjadi tabung oral telah ditemukan sebagai sinapomorfi kelompok tersebut (Faivovich et al. 2005 ; Orrico et al. 2021 ). Selain kesamaan yang dibahas dalam literatur, terdapat beberapa perbedaan dalam konfigurasi tabung oral di antara beberapa spesies. Sementara pada sebagian besar spesies labium bawah terlipat menjadi bentuk U (misalnya, D nanus, D. branneri, D. leali ; lihat Gambar 9 , hal. 218, Vera-Candioti et al. 2004 ; Schulze et al. 2015 ; De Abreu et al. 2015 ), pada D. cerradensis dan D. mathiassoni lipatan eksternal menutupi hampir seluruh labia (Kaplan 2017 ; penelitian ini). Pada D. microcephalus dan D. rhodopeplus lipatan luar yang menutupi labium atas pendek dan berkerut, dengan labium atas dan tonjolan atas terlihat, sedangkan pada D. cerradensis dan D. mathiassoni lipatan luarnya panjang dan halus, menutupi labium atas dan hampir seluruh labium bawah. Karena kondisi ini, lubang mulut mungkin lebih berposisi ventral pada kedua spesies, dan hanya tepi labium bawah dan tonjolan atas yang terlihat.
Kelompok Dendropsophus microcephalus (yaitu, D. microcephalus, D. nanus , D. phlebodes ; Wassersug 1980 ; Vera-Candioti 2007 ) menunjukkan pengurangan elemen buccopharyngeal, suatu kondisi yang terkait dengan makrofagi. Sebaliknya, kelompok spesies Dendropsophus decipiens, D. parviceps, D. marmoratus , D. minutus , dan D. leucophyllatus memiliki lebih banyak elemen di dasar dan atap bukal (misalnya, papila, medial ridge, papila lateral ridge; Echeverría 1997 ; Kaplan dan Ruiz 1997 ; Wassersug 1980 ; Vera-Candioti 2007 ; Dias et al. 2019 ; Costa et al. 2024 ). Kelompok-kelompok ini (kecuali kelompok D. leucophyllatus ), serta Xenohyla truncata (Dias et al. 2023 ), memiliki papila berbentuk kipas yang membatasi dasar bukal (Wassersug 1980 ; Dias et al. 2019 ). Dengan mempertimbangkan hubungan filogenetik Dendropsophini (Orrico et al. 2021 ; Whitcher et al. 2025 ), ada kemungkinan bahwa hilangnya papila ini telah berevolusi pada nenek moyang bersama kelompok D. leucophyllatus + D. microcephalus .
Kehadiran papila prenarial yang menutupi nares internal di D. cerradensis dilaporkan untuk pertama kalinya di suku Dendropsophini. Seperti D. cerradensis , D. microcephalus dan D. nanus juga memiliki nares internal yang diposisikan pada sudut lancip relatif terhadap sumbu longitudinal, meskipun papila prenarial belum dijelaskan untuk spesies ini (Vera-Candioti 2007 ). Namun, proyeksi terlihat di nares internal D. nanus (lihat gbr. 29, dalam Vera-Candioti 2007 : 59), dan fitur ini dapat mewakili karakter turunan dalam kelompok tersebut. Karena papila menutupi nares internal, mereka mungkin memiliki fungsi perlindungan mekanis saat makanan ditelan oleh isapan (lihat hipotesis serupa di Wassersug 1980 : 113).
Dendropsophus cerradensis dan spesies lain dari kelompok D. microcephalus memiliki papila infralabial bulat (Wassersug 1980 ; Vera-Candioti 2007 ). Telah dikemukakan bahwa kecebong menggunakan papila infralabial mereka untuk mendeteksi posisi makanan nabati (Wassersug 1980 ). Dalam hal ini, ada kemungkinan bahwa kelompok D. microcephalus menggunakan sarung rahang mereka untuk menggigit atau memegang makanan, seperti yang diamati pada D. cerradensis , berbeda dengan anura lain, yang menggunakan rahang mereka terutama untuk mengikis partikel makanan dari substrat.
Suprarostral sebagai elemen tunggal adalah sinapomorfi putatif dari Dendropsophini (kecuali dalam D. ebraccatus ; Haas 2003 ; Dias et al. 2023 ). Jika tidak, tidak adanya prosesus anteroposterior dalam suprarostral mungkin merupakan sinapomorfi dari kelompok D. microcephalus (yaitu, prosesus anteroposterior hadir dalam D. decipiens , D. molitor , dan D. soaresi , dengan yang terakhir menunjukkan takik berbentuk V medial kecil). Fitur lain mungkin juga eksklusif untuk kelompok D. microcephalus , seperti prosesus muskular, yang berbentuk segi empat dalam kelompok ini, tetapi berbentuk segitiga dalam D. decipiens, D. molitor , dan D. soaresi (Dias et al. 2019 ; Arenas-Rodríguez et al. 2018 ; Costa et al. 2024 ). M. subarcualis rectus II–IV terputus pada D. cerradensis , D. microcephalus , dan D. nanus (Vera-Candioti 2007 ), sementara itu merupakan slip terus menerus pada berudu Dendropsophini lainnya, seperti Xenohyla truncata , D. decipiens , dan D. ebraccatus (Haas 2003 ; Dias dkk. 2019 , 2023 ).
Insersi m. levator mandibulae longus profundus pada kartilago Meckel, seperti yang diamati pada D. cerradensis , merupakan kejadian langka pada Neobatrachia, karena konfigurasi ini hanya dideskripsikan untuk Rhacophorus vampyrus (Vera-Candioti et al. 2021 ). Pada spesies dengan satu slip mlm longus, seperti Ascaphus truei (Ascaphidae), Rhinophrynus dorsalis (Rhinophrynidae), Pipa carvalhoi (Pipidae), insersi terjadi pada rahang bawah (Haas 2003 ). Sebaliknya, pada spesies dengan mlm longus dalam dua bagian (yaitu, superfisialis dan profundus), bagian m. levator mandibulae longus profundus menyisipkan pada kartilago suprarostral, yang merupakan kondisi umum pada Anura (misalnya, Haas 2003 ; Vera-Candioti 2007 ).
Meskipun perilaku makan kelompok ini telah dibahas dalam literatur (yaitu, Lavilla 1990 ; Vera-Candioti et al. 2004 ; Vera-Candioti 2007 ; Kaplan 2017 ), tidak ada rekaman video sampai penelitian kami. Video menunjukkan penonjolan tabung oral yang lengkap dan cepat, yang menunjukkan mekanisme makan hisap yang efisien. Perilaku ini juga dapat terjadi pada spesies lain dari kelompok D. microcephalus , tetapi perbedaan perilaku dan/atau mekanis dapat terjadi karena perbedaan tingkat internalisasi tabung oral (lihat di atas). Data kami menguatkan bahwa mr ab. anterior dimasukkan ke ujung paling anterior lipatan eksternal (Kaplan 2017 ), kondisi unik di Anura. Insersi anterior mr ab. telah dilaporkan pada kecebong penghisap dan kecebong rheophilous (misalnya, Noble dan Pope 1929 ; Haas dan Richards 1998 ; Dias et al. 2024 ), namun tidak pernah menyisipkan pada jaringan lunak.
Meskipun kami masih belum dapat menyelesaikan sepenuhnya mekanisme penonjolan tabung oral, pengamatan kami mendukung hipotesis penonjolan tabung oral (Lavilla 1990 ; Vera-Candioti et al. 2004 ; Vera-Candioti 2007 ). Namun, ada kemungkinan juga bahwa lipatan eksternal ditarik oleh kontraksi m. rectus abdominis anterior, yang berkontribusi pada paparan tabung oral, seperti yang disarankan oleh Kaplan ( 2017 ). Dengan demikian, perilaku makan hisap kelompok D. microcephalus dapat disebabkan oleh mekanisme ganda yang melibatkan penarikan lipatan eksternal dan penonjolan tabung oral itu sendiri secara bersamaan atau selanjutnya. Selain itu, m. mandibulolabialis dapat berperan dalam kontrol motorik halus tabung oral, karena mungkin terlibat, setidaknya dalam penarikan tabung (lihat Vera-Candioti et al. 2004 ). Pada spesies dengan baris gigi, m. mandibulolabialis biasanya berinsersi pada tonjolan dan mungkin berfungsi untuk mendistorsi tonjolan tersebut secara distal (Carr dan Altig 1991 ; tetapi lihat Dias 2020 ). Investigasi lebih lanjut terhadap otot-otot ini, misalnya menggunakan pendekatan elektromiografi, dapat menjelaskan fungsinya.
Makan dengan cara menghisap adalah strategi yang ditemukan di banyak kelompok vertebrata (misalnya, Deban dan Olson 2002 ; Kleinteich et al. 2014 ; Bellwood et al. 2015 ). Di antara beberapa strategi dan adaptasi yang terkait dengan cara makan ini, penjuluran rahang adalah salah satu inovasi terpenting dalam makan vertebrata (Wainwright et al. 2015 ). Di dalam amfibi, makan dengan cara menghisap digunakan oleh salamander akuatik (baik dewasa maupun larva), larva caecilian, dan beberapa anuran (misalnya, O’Reilly 2000 ; Deban 2003 ; Carreño dan Nishikawa 2010 ; Barrionuevo 2016 ; Duport-Bru dan Abdala 2024 ).
Mengenai anuran, makan dengan cara menghisap diamati pada spesies akuatik, seperti Pipidae (Duellman dan Trueb 1994 ), tetapi kebanyakan kecebong terutama makan dengan cara mengikis dan/atau menyaring. Meskipun menghisap adalah perilaku makan leluhur vertebrata, termasuk kecebong, D. cerradensis dan mungkin spesies lain dari kelompok D. microcephalus menunjukkan sebuah inovasi: sebuah tabung mulut yang tetap ditarik dan hanya menonjol pada saat dihisap. Occidozyga baluensis (Dicroglossidae) adalah satu-satunya spesies dengan tabung mulut yang sama, berbagi fitur lain, seperti selubung rahang yang kuat dan ceratohyal, dan keranjang branchial yang berkurang (Haas et al. 2014 ; Vassilieva et al. 2025 ). Namun, dalam O. baluensis , m. rectus abdominis anterior menyisipkan dari diafragma ke kulit ventral. Kehadiran cacing utuh atau serangga mirip cacing di dalam isi ususnya memberikan bukti adanya makan dengan cara menghisap, walaupun penonjolan tabung mulut belum terdokumentasikan pada spesies ini (Haas et al. 2014 ; Vassilieva et al. 2025 ).
Hymenochirus boettgeri (Pipidae) adalah kecebong makrofag lain yang memanfaatkan makan dengan cara menghisap. Cakram oral H. boettgeri adalah tabung eksternal yang terlipat ke arah anterodorsal dalam posisi istirahat dan “diregangkan” selama perilaku makan (lihat Gambar 2 , hlm. 275, Sokol 1962 ). Meskipun durasi peristiwa menghisap pada D. cerradensis tidak dihitung secara tepat, durasinya sama dengan atau kurang dari 0,016 detik, mirip dengan yang dilaporkan untuk H. boettgeri (0,04–0,012 detik), yang lebih cepat daripada larva teleost (Deban dan Olson 2002 ). Mengingat bahwa ukuran lubang mulut berbanding terbalik dengan tekanan yang dihasilkan (Wassersug dan Hoff 1979 ), isapan yang dicetak oleh tabung mulut kecil dan kekokohan otot-otot hyoid di D. cerradensis , mungkin menghasilkan daya isap yang besar, yang memungkinkan pemangsaan seluruh oligochaetes yang tersembunyi di akar tanaman atau di substrat. Apakah ada fleksibilitas makanan pada kecebong kelompok ini, yang memakan cacing dan bahan tanaman, atau apakah tanaman merupakan komponen sekunder yang tertelan, masih belum diketahui dan memerlukan penyelidikan lebih lanjut (Wassersug dan Rosenberg 1979 ; Vera-Candioti et al. 2004 ; Vera-Candioti 2007 ).
Perbedaan lain antara ketiga garis keturunan yang tidak berhubungan ini berkenaan dengan mekanisme lokalisasi mangsa. Baik H. boettgeri dan O. baluensis memiliki mata anterior dengan penglihatan stereoskopis (Deban dan Olson 2002 ; Haas et al. 2014 ), yang mungkin berperan dalam mendeteksi makanan, sementara D. cerradensis memiliki mata dorsolateral dan mungkin bergantung pada indera lain untuk “berburu”. Dendropsophus cerradensis memiliki kantung hidung yang berkembang dengan baik yang berfungsi sebagai titik perlekatan pada lm lateralis. Seperti yang disarankan oleh Haas ( 2003 ), kontraksi otot ini dapat melebarkan kantung hidung, yang berpotensi meningkatkan fungsi penciumannya.
5 Kesimpulan
Perilaku hisap yang dilaporkan di sini untuk berudu D. cerradensis menunjukkan mode makan yang unik di Anura, yang dicirikan oleh tonjolan penuh tabung mulut secara eksklusif selama makan. Dendropsophus cerradensis memiliki tabung mulut yang sepenuhnya terinternalisasi, sementara morfologi eksternal dan internal yang tersisa mirip dengan spesies lain yang diketahui dari kelompok D. microcephalus , yang menunjukkan adaptasi terhadap makrofagi. Fitur utama dari adaptasi ini meliputi: (1) cakram mulut dimodifikasi dalam tabung mulut, (2) tabung mulut diposisikan secara anteroventral, (3) tubuh memanjang dan tertekan, (4) posisi lubang hidung di ujung moncong, (5) perkembangan kapsul hidung, (6) elemen berkurang dalam rongga bukofaring, (7) orbitohyoideus sebagai otot utama yang kuat, (8) insersi m. rectus abdominis superfisialis di lipatan eksternal, (9) ceratohyal yang kuat dan melengkung, (10) keranjang branchial yang berkurang. Meskipun beberapa aspek perilaku diamati, seperti membungkukkan badan untuk mencari makanan di substrat, sulit untuk menarik kesimpulan tentang perilaku predator lain dari D. cerradensis . Masih belum diketahui apakah kecebong ini adalah predator aktif atau bergantung pada strategi duduk dan menunggu. Meskipun konvergensi diamati dalam rongga bukafaring dan sistem muskuloskeletal dengan keluarga anuran lainnya, tabung mulut yang diinternalisasi tetap menjadi fitur unik dari kelompok D. microcephalus . Penelitian lebih lanjut sangat penting untuk mengetahui apakah spesialisasi ini dapat mewakili kebaruan evolusi yang memfasilitasi pendudukan zona adaptif baru dan eksploitasi sumber daya baru.