
ABSTRAK
Deteksi gerakan merupakan aspek dasar dari sebagian besar sistem visual hewan. Akan tetapi, banyak lingkungan yang rentan terhadap gerakan latar belakang, yang dapat mengganggu kemampuan sistem visual untuk mendeteksi isyarat gerakan yang relevan. Sementara pada manusia, gerakan latar belakang dapat mengganggu deteksi isyarat visual bahkan setelah komponen latar belakang yang bergerak telah berhenti, masih belum diketahui apakah bentuk alami gerakan latar belakang juga dapat memengaruhi sistem visual hewan lainnya. Di sini, kami menguji apakah paparan sebelumnya terhadap ‘bahan kimia’ yang terjadi secara alami, suatu bentuk pola cahaya yang bergerak secara dinamis yang umum ditemukan di lingkungan perairan dangkal, dapat memiliki efek yang bertahan lama pada kemampuan deteksi gerakan hewan bahkan setelah paparan bahan kimia tersebut berhenti. Untuk melakukan ini, kami menetapkan probabilitas respons kepiting pantai Carcinus maenas terhadap rangsangan cakram mengembang yang dihasilkan komputer yang meniru predator yang mendekat setelah paparan terhadap pemandangan bahan kimia statis atau bergerak. Paparan sebelumnya terhadap bahan kimia yang bergerak memiliki efek bertahan jangka pendek pada persepsi visual pada C. maenas , mengurangi kemungkinan kepiting untuk merespons predator yang mendekat setidaknya selama 2 detik setelah bahan kimia yang bergerak berhenti. Studi kami menunjukkan bahwa bahkan setelah periode paparan terhadap gerakan latar belakang berakhir, tingkat respons visual pada C. maenas masih dapat berkurang untuk waktu yang singkat karena paparan sebelumnya. Sementara apa yang disebut ‘efek historis’ ini mungkin berasal dari adaptasi sistem visual kepiting terhadap gerakan latar belakang yang keras, kami membahas apakah hal itu dapat berdampak pada kelangsungan hidup spesies krustasea ini.
1 Pendahuluan
Banyak hewan mengandalkan sistem visual mereka untuk mendeteksi informasi tentang predator, mangsa, atau sesama spesies. Meskipun mata hewan dapat bervariasi baik dalam anatomi maupun kompleksitasnya, beberapa fungsi umum di antara sistem visual, menyoroti pentingnya ekologisnya bagi kelangsungan hidup dan keberhasilan reproduksi hewan (Land dan Nilsson 2012 ). Salah satu fungsi ini adalah kemampuan untuk mendeteksi gerakan, yang digunakan untuk berbagai proses perilaku seperti navigasi, stabilitas postur, penangkapan mangsa atau deteksi predator (Eckert dan Zeil 2001 ; Srinivasan et al. 1999 ). Namun, banyak lingkungan alami rentan terhadap gerakan latar belakang, misalnya melalui vegetasi yang tertiup angin atau pergerakan awan atau permukaan air, seperti sungai atau air terjun, yang menciptakan elemen visual dinamis di seluruh bagian pemandangan visual yang terkadang besar. Gerakan latar belakang tersebut dapat bertindak sebagai sumber gangguan visual, yang mengganggu kemampuan hewan untuk mendeteksi dan menanggapi isyarat gerakan (Attwell et al. 2021 ; Matchette et al. 2020 ; Venables et al. 2022 ). Secara khusus, gerakan latar belakang dapat mengganggu persepsi hewan melalui dua proses, baik dengan menutupi rangsangan bergerak lainnya dalam pemandangan (Matchette et al. 2018 , 2019 ) atau dengan mengalihkan perhatian hewan, sehingga membatasi kemampuannya untuk mendeteksi, menanggapi, atau memproses informasi visual (Dominoni et al. 2020 ).
Meskipun gerakan latar belakang dapat mengganggu banyak proses perilaku dan persepsi pada hewan, penelitian sebelumnya belum mengungkap apakah dampak ini berasal secara eksklusif dari gangguan yang dapat diciptakan oleh gerakan latar belakang pada saat deteksi isyarat (terlepas dari paparan sebelumnya terhadap pemandangan latar belakang yang bergerak) atau apakah paparan sebelumnya terhadap gerakan latar belakang juga memengaruhi pemrosesan visual hewan. Misalnya, sistem visual dapat melakukan adaptasi saraf terhadap gerakan latar belakang yang berkelanjutan, sehingga meningkatkan kemungkinan hewan untuk mendeteksi rangsangan bergerak lainnya (Webster 2015 ). Secara khusus, karena adaptasi, individu yang mengalami paparan yang lama terhadap latar belakang yang bergerak mungkin lebih cenderung mendeteksi dan merespons isyarat yang bergerak (misalnya, predator yang bergerak) terhadap latar belakang yang bergerak daripada individu yang memiliki lebih sedikit waktu untuk beradaptasi dengan latar belakang tersebut. Namun, paparan sebelumnya terhadap gerakan latar belakang juga dapat memengaruhi sistem visual hewan setelah gerakan latar belakang berhenti. Pada manusia, misalnya, mengalihkan pandangan dari pemandangan dengan paparan berkelanjutan terhadap stimulus bergerak (misalnya, air terjun) ke latar belakang yang diam dapat menciptakan ilusi objek diam yang bergerak ke arah yang berlawanan dengan stimulus bergerak sebelumnya (Anstis et al. 1998 ), sebuah fenomena yang dikenal sebagai ‘ilusi air terjun’ (Addams 1834 ). ‘Efek historis’ dari gerakan latar belakang ini dapat mengganggu deteksi informasi, sehingga berpotensi mengganggu proses persepsi dan pengambilan keputusan (Gallagher et al. 2021 ). Melihat di luar persepsi manusia, efek historis gerakan latar belakang tersebut juga telah ditetapkan pada primata; namun, penelitian ini memaparkan hewan pada stimulus bergerak yang tidak alami, seperti kisi gelombang sinus yang melayang (Glasser et al. 2011 ; Kohn dan Movshon 2003 ). Apakah gerakan latar belakang yang terjadi secara alami dan relevan secara ekologis, seperti pola cahaya bergerak yang berasal dari pencahayaan dinamis, dapat menyebabkan efek historis pada sistem visual hewan masih belum diketahui.
Salah satu bentuk umum gerakan latar belakang di habitat perairan dangkal adalah ‘kaustik’ (juga disebut kedipan kaustik; Gambar 1a,b ). Terdiri dari pola seperti jaring dari pita cahaya intensitas tinggi yang bergerak melintasi substrat, kaustik dihasilkan dari fluktuasi titik fokus berkas cahaya saat dibiaskan oleh gelombang permukaan (Lock dan Andrews 1992 ). Di lingkungan yang rentan terhadap pencahayaan kaustik, hewan mungkin menggunakan isyarat gerakan nyata yang disediakan oleh bentuk penerangan dinamis ini untuk menutupi gerakan mereka sendiri, yang pada gilirannya dapat meningkatkan kesulitan bagi predator atau mangsa untuk mendeteksinya (Matchette et al. 2018 ). Memang, paparan bahan kimia kaustik dapat mengganggu deteksi mangsa atau predator pada ikan picu Picasso ( Rhinecanthus aculeatus ; Matchette et al. 2020 ), ikan stickleback berduri tiga ( Gasterosteus aculeatus ; Attwell et al. 2021 ), kepiting pantai ( Carcinus maenas ; Venables et al. 2022 ) dan sotong Eropa ( Sepia officinalis ; Venables et al. 2022 ). Meskipun pola cahaya kaustik yang bergerak secara dinamis dapat mengganggu sistem visual hewan akuatik (Attwell et al. 2021 ; Matchette et al. 2020 , tetapi lihat Drerup et al. 2023 ; Drerup, How, et al. 2024 ; Venables et al. 2022 ), masih belum diketahui apakah hanya gerakan latar belakang pada saat penyajian isyarat (terlepas dari paparan kaustik sebelumnya) yang mengurangi kemampuan untuk merespons objek-objek ini pada spesies ini, atau apakah paparan kaustik sebelumnya dapat meningkatkan atau lebih lanjut mengganggu persepsi visual hewan-hewan ini. Memang, kaustik dapat muncul dan menghilang secara berurutan dengan cepat karena tutupan awan atau kondisi angin yang berfluktuasi, yang memaparkan hewan di lingkungan laut dangkal pada fase gerakan latar belakang yang tidak dapat diprediksi. Oleh karena itu, kaustik merupakan bentuk gerakan latar belakang yang terjadi secara alami yang berpotensi menimbulkan efek historis pada sistem visual hewan laut.
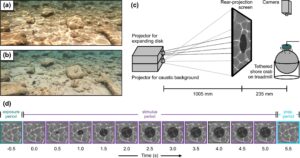
Dalam studi ini, kami menyelidiki efek temporal gerakan latar belakang pada kemampuan kepiting pantai ( C. maenas ) untuk mendeteksi predator. Spesies kepiting brachyuran ini umumnya ditemukan di lingkungan pesisir seperti zona pasang surut dan subpasang surut dangkal (Crothers 1968 ) di mana ia mengandalkan sistem visualnya, yang terdiri dari sepasang mata majemuk, untuk banyak tugas perilaku, termasuk mendeteksi predator, menangkap mangsa, dan menemukan spesies yang sama (Cronin dan Feller 2014 ). Dengan demikian, mata mereka disetel untuk mendeteksi berbagai bentuk gerakan di bidang visual (Horseman et al. 2011 ). Mempertimbangkan bahwa habitat alami mereka rentan terhadap pola pencahayaan kaustik yang muncul dan menghilang dengan cepat yang dapat mengganggu persepsi visual spesies ini (Venables et al. 2022 ), C. maenas menawarkan model yang cocok untuk menetapkan efek temporal gerakan latar belakang yang disebabkan oleh pencahayaan dinamis pada kemampuan untuk mendeteksi rangsangan yang bergerak. Dengan memaparkan individu-individu C. maenas pada perubahan pemandangan visual yang terdiri dari simulasi buatan komputer berupa pola-pola kaustik yang bergerak maupun statis, sekaligus memperlihatkan stimulus yang muncul menirukan predator yang mendekat, kami menilai apakah paparan terhadap kaustik yang bergerak dapat menimbulkan dampak historis yang relevan secara ekologis terhadap deteksi gerakan pada kepiting.
2 Bahan dan Metode
2.1 Organisme yang diteliti
Kepiting pantai C. maenas dikumpulkan dari Pantai Clevedon (Inggris; 51°26′18.0″ N 2°51′56.7″ W) dan dipelihara dalam kompartemen individual (180 × 130 × 60 mm) dalam akuarium dangkal bersirkulasi yang diisi dengan air laut buatan (Tropic Marin AG, Wartenberg, Jerman) pada salinitas 35 ppt. Semua kompartemen penampungan terkena siklus cahaya yang sesuai dengan fotoperiode alami periode percobaan (Mei–Juni). Kepiting pantai diberi makan dua kali seminggu dengan kerang atau udang yang dicairkan. Semua individu dikembalikan ke lokasi pengumpulan dalam waktu seminggu setelah pengumpulan.
Karena kami menggunakan individu C. maenas yang ditangkap di alam liar , kami tidak dapat secara langsung mengendalikan perbedaan individu dalam pengalaman sebelumnya terhadap paparan cahaya kaustik. Namun, penelitian kami tidak bertujuan untuk menilai respons C. maenas terhadap rangsangan visual baru, melainkan untuk menetapkan bagaimana sistem visual C. maenas mengatasi gerakan latar belakang yang terjadi secara alami di habitatnya, yang menghilangkan kebutuhan untuk menguji individu yang naif. Dengan mengingat hal ini, kami bertujuan untuk menjaga efek pengalaman individu seminimal mungkin dengan hanya mengumpulkan individu dengan ukuran yang sebanding dari lokasi geografis yang sama (segmen pantai selebar 200 m) dalam jangka waktu yang singkat (3 minggu), sementara lebih lanjut mengendalikan variasi individu dengan memasukkan ID individu sebagai efek acak dalam model statistik kami (lihat Bagian 2.8 , Analisis Statistik).
2.2 Pengaturan Eksperimen
Kepiting diikat di atas treadmill berbentuk bola yang terdiri dari bola Styrofoam (diameter = 120 mm) yang digantung di atas bantalan udara yang disuplai dari keran udara bertekanan (Gambar 1c ). Penambat melibatkan perekatan sepotong Velcro ke sisi dorsal karapas kepiting menggunakan lem sianoakrilat dan sepotong Velcro pelengkap ke batang logam yang dipasang secara horizontal di atas treadmill. Ini memungkinkan kepiting berjalan bebas tetapi membatasi gerakan translasi atau rotasi mereka. Sebuah kamera video (HC-X900, Panasonic Corporation, Osaka, Jepang) dipasang 375 mm di atas treadmill untuk merekam perilaku kepiting di setiap percobaan.
Kepiting menghadapi layar stimulus yang dibuat khusus pada jarak 235 mm yang memungkinkan tampilan latar belakang kaustik dan stimulus predator yang mendekat secara bersamaan namun independen. Layar ini terdiri dari dua proyektor digital (PA503S, ViewSonic Corporation, Brea, AS) yang ditumpuk di atas satu sama lain, dengan gambar mereka melapisi layar proyeksi belakang 300 × 300 mm semi-transparan (0,5 diffusor, Lee Filters, Andover, Inggris) pada jarak 1005 mm (Gambar 1c ). Satu proyektor, yang terhubung ke laptop (Ideapad 310, Lenovo Group Limited, Hong Kong, VR China), menampilkan animasi yang dihasilkan komputer dari pola kaustik yang dirender menggunakan Caustics Generator Pro (Dual Heights; www.dualheights.se/caustics/ ; pengaturan perangkat lunak terperinci disajikan dalam Tabel A1 ). Pola kaustik terdiri dari 200 bingkai unik yang terus-menerus diputar pada laju bingkai 30 bingkai/detik, menghasilkan durasi putaran tunggal selama 6,66 detik. Proyektor kedua, yang terhubung ke laptop kedua (G3, Dell Technologies Inc., Round Rock, AS), digunakan untuk memproyeksikan stimulus cakram hitam yang mengembang yang dihasilkan menggunakan skrip MATLAB yang dibuat khusus (R2021a, MathWorks, Natick, AS). Stimulus cakram hitam yang mengembang merupakan pendekatan eksperimental umum untuk meniru kemunculan predator yang mendekat (Calanni et al. 2024 ; Schiff et al. 1962 ), yang menimbulkan respons beku bawaan yang kuat pada C. maenas (Drerup dan How 2021 ; Venables et al. 2022 ; lihat 2.7 , Penilaian respons). Cakram hitam mengembang selama periode 2 detik dari sudut pandang 0° hingga ~33° (0–139 mm diameter), mengikuti profil ekspansi geometris (yaitu, laju sudut ekspansi cakram cocok dengan yang diinduksi oleh objek fisik yang mendekat dengan kecepatan konstan), dan tetap terlihat pada ekspansi penuh selama 3 detik berikutnya (Gambar 1d ). Karena kepiting ‘berwajah lebar’ sebagian besar tidak memiliki zona akut di mata majemuknya (Zeil et al. 1986 ), set-up kami memaparkan individu C. maenas yang ditambatkan dengan stimulus cakram yang mengembang di ekuator mata mereka, dengan stimulus seperti itu di bagian bidang pandang ini telah terbukti menimbulkan respons antipredator yang kuat (misalnya, Drerup dan How 2021 ; Venables et al. 2022 ). Kedua proyektor dijalankan pada kecepatan refresh 60 Hz, yang berada di atas frekuensi fusi kedipan kritis kepiting (sekitar 30–50 Hz; Grober 1990 ; Layne et al. 1997). Bunyi bip audio yang dihasilkan oleh skrip MATLAB di awal dan akhir setiap presentasi cakram yang mengembang dimasukkan ke aliran audio kamera video untuk menyinkronkan respons kepiting terhadap stimulus. Untuk meminimalkan gangguan eksternal, bilik putih yang terdiri dari tenda fotografi selebar 40 × 40 × 40 cm ditempatkan di sekitar treadmill kepiting, dengan dua bukaan yang memungkinkan kepiting menghadap layar proyeksi belakang, serta kamera untuk merekam perilaku kepiting.
Menggunakan set-up kami (terdiri dari dua proyektor dan melapisi gambar mereka ke layar proyeksi belakang) memiliki tiga keuntungan. Pertama, menggunakan dua aliran animasi independen untuk menampilkan kaustik melalui satu proyektor dan stimulus cakram yang mengembang melalui proyektor lainnya memungkinkan kami untuk memulai/menghentikan animasi secara independen satu sama lain. Ini memungkinkan kami untuk mengatur waktu presentasi stimulus ke fase-fase di mana kepiting pantai bergerak, yang berkaitan dengan pendekatan penilaian respons kami (lihat 2.7 , Penilaian Respons). Kedua, penggunaan proyektor pemrosesan cahaya digital (DLP) dalam set-up kami memungkinkan kami untuk menampilkan latar belakang kaustik serta stimulus cakram yang mengembang sebagai isyarat berbasis intensitas saja, tanpa fluktuasi atau artefak yang tidak diinginkan dalam polarisasi cahaya. C. maenas mampu mengurangi dampak pencahayaan kaustik pada kemampuannya mendeteksi objek jika objek tersebut terpolarisasi (Venables et al. 2022 ), sehingga pengaturan eksperimen kami memastikan bahwa respons perilaku apa pun yang diamati dalam penelitian ini semata-mata ditimbulkan oleh dampak kaustik pada saluran visual berbasis intensitas spesies ini. Ketiga, dengan memproyeksikan animasi kaustik dan stimulus cakram yang mengembang ke layar proyeksi belakang, kedua presentasi tersebut tampak bagi kepiting pada bidang visual yang sama. Pendekatan ini memungkinkan kami untuk melapiskan kaustik ke latar belakang maupun stimulus cakram yang mengembang, sehingga memperkirakan bagaimana kaustik alami akan berinteraksi dengan objek yang mendekat (misalnya, predator) dengan latar belakang. Oleh karena itu, desain eksperimen kami menggambarkan skenario di mana seekor kepiting didekati oleh predator yang dilihat dengan latar belakang vertikal atau miring, seperti yang mungkin ditemukan di habitat pesisir dangkal atau kolam batu besar. Meskipun pengaturan kami menghasilkan gerakan dalam bentuk pola cahaya kaustik yang bergerak dinamis di latar belakang visual dan latar depan, demi kesederhanaan kami menyebut perlakuan kaustik kami sebagai gerakan latar belakang di seluruh sisa naskah.
2.3 Pengukuran Kontras
Perlakuan kami terdiri dari animasi kaustik yang dilapisi oleh stimulus mirip predator yang mendekat yang terdiri dari cakram hitam (nilai uint8 = 0) yang mengembang pada latar belakang dengan nilai skala abu-abu yang sama atau lebih terang (nilai uint8 antara 0 dan 255). Untuk menghitung kontras antara stimulus mirip predator (cakram mengembang yang dilapisi oleh kaustik) dan latar belakang (latar belakang skala abu-abu yang dilapisi oleh kaustik), pertama-tama kami memproyeksikan bingkai diam animasi kaustik (menggunakan proyektor pertama) serta serangkaian 256 bingkai yang terdiri dari semua 256 nilai skala abu-abu uint8 (menggunakan proyektor kedua) ke layar proyeksi belakang. Bahasa Indonesia: Dengan menggunakan spektrometer (HDX) yang digandengkan ke serat optik polos 400 μm dengan korektor kosinus (R400-7-UV-VIS; semua Ocean Insight, Orlando, AS), kami kemudian mengukur radiansi dari subset gambar yang dihamparkan, yang mencakup sekitar setengah dari area layar, untuk semua kombinasi latar belakang kaustik dan masing-masing dari 256 nilai skala abu-abu. Pengukuran radiansi dibatasi pada rentang panjang gelombang 400–700 nm, yang mencakup sensitivitas spektral C. maenas (Bruno et al. 1973 ; Wald 1968 ). Kami kemudian menggunakan pengukuran radiansi ini untuk menghitung kontras Weber dari semua 256 nilai skala abu-abu (Gambar A1a ), yang digunakan untuk latar belakang proyeksi stimulus, terhadap gambar skala abu-abu paling gelap (nilai uint8 = 0), yang digunakan untuk cakram yang mengembang dari proyeksi stimulus, menggunakan rumus berikut:
![]()
Untuk menentukan rentang kontras stimulus-latar belakang eksperimental yang sesuai, kami melakukan eksperimen percontohan yang menguji tingkat respons 11 individu C. maenas (tidak digunakan kembali dalam Eksperimen 1 atau 2) terhadap 11 cakram yang mengembang dengan kontras Weber antara -0,10 dan -0,27 terhadap latar belakang kaustik statis, yang disajikan dalam desain blok acak untuk menghindari bias urutan. Respons kepiting pantai terhadap stimulus yang muncul ditetapkan menggunakan sistem penilaian biner (lihat Bagian 2.7 , Penilaian Respons) dan ditampilkan dalam Gambar A1b .
2.4 Percobaan 1
Untuk menetapkan efek temporal gerakan latar belakang pada respons C. maenas , kami memaparkan 56 individu (lebar karapas: 32 ± 9 mm [rata-rata ± 1 SD]) masing-masing pada delapan perlakuan berbeda. Setiap perlakuan dimulai dengan ‘periode pemaparan’ ≥ 90 detik pada latar belakang kaustik statis atau bergerak dan segera diikuti oleh ‘periode stimulus’, yang bisa jadi merupakan kelanjutan dari kondisi kaustik yang sama atau perubahan mendadak ke kondisi kaustik lainnya. Dalam periode stimulus, kepiting kemudian terpapar pada stimulus cakram yang mengembang, yang dimulai bersamaan dengan perubahan dari periode pemaparan ke periode stimulus, sehingga mencapai ukuran penuhnya 2 detik dalam periode stimulus (Gambar 1d ). Dalam empat dari delapan perlakuan percobaan ini, cakram yang mengembang diproyeksikan pada kontras Weber sebesar -0,22 (Gambar A1b ), yang dianggap sebagai kontras terendah yang memunculkan tingkat respons yang kuat (~90%) terhadap cakram yang mengembang terhadap latar belakang kaustik statis tetapi dengan demikian tingkat respons yang lebih rendah terhadap kaustik yang bergerak (mengikuti Venables et al. 2022 ). Dalam empat perlakuan lainnya, cakram yang mengembang disajikan pada kontras Weber sebesar 0,00, yang menghasilkan cakram yang mengembang ‘tak terlihat’ dan digunakan sebagai kontrol untuk menetapkan apakah ada respons perilaku yang diamati yang benar-benar berasal dari paparan rangsangan seperti predator yang mendekat dan bukan dari perubahan potensial pada latar belakang visual, dengan demikian menilai proporsi respons positif palsu. Kami memilih untuk menyajikan kepiting dengan cakram tak kasat mata alih-alih tanpa cakram sama sekali untuk memastikan bahwa respons yang diamati dalam perilaku kepiting tidak disebabkan oleh penyimpangan dalam protokol eksperimen kami (misalnya, permulaan manual tampilan cakram yang mengembang oleh peneliti atau gangguan apa pun dalam animasi kaustik yang disebabkan oleh penyajian cakram yang mengembang). Secara total, ada delapan perlakuan, dengan variasi kaustik statis atau bergerak baik dalam periode paparan maupun stimulus, serta kontras (‘terlihat’ atau ‘tak terlihat’) dari cakram yang mengembang. Kami memberi setiap perlakuan kode tiga huruf, dengan huruf pertama dan kedua menyatakan kondisi kaustik dalam paparan dan kemudian periode stimulus (‘M’ untuk kaustik bergerak, ‘S’ untuk kaustik statis), dan huruf ketiga menyatakan visibilitas cakram yang mengembang (‘V’ untuk cakram yang terlihat dengan kontras Weber -0,22, ‘I’ untuk cakram tak terlihat dengan kontras Weber 0,00). Setiap kepiting ( n = 56) menerima semua delapan perlakuan dalam satu percobaan dan hanya diuji sekali. Untuk memastikan bahwa urutan perlakuan yang diberikan kepada individu dalam suatu percobaan tidak memengaruhi kemungkinan responsnya, kami membuat 56 urutan unik dari delapan perlakuan dalam suatu rancangan blok acak seimbang, dengan demikian setiap kepiting menerima urutan unik dari delapan perlakuan dalam percobaannya.
2.5 Percobaan 2
Berdasarkan hasil Percobaan 1, kami menentukan berapa lama paparan sebelumnya terhadap kaustik yang bergerak mengurangi tingkat respons terhadap rangsangan yang muncul setelah kaustik berhenti. Untuk melakukannya, kami memaparkan 48 kepiting baru (yaitu, tidak digunakan dalam Percobaan 1; lebar karapas: 31 ± 8 mm [rata-rata ± 1 SD]) pada delapan perlakuan yang berbeda. Setiap perlakuan dimulai dengan ‘periode paparan’ ≥ 90 detik pada latar belakang kaustik statis atau bergerak dan segera diikuti oleh ‘periode stimulus’, yang dalam percobaan ini selalu terdiri dari latar belakang kaustik statis. Pada periode stimulus, kami kemudian menyajikan cakram yang mengembang dengan titik awal yang berbeda, yaitu setelah 0 detik (dengan demikian dimulai dengan perubahan dari paparan ke periode stimulus) atau dengan offset 1, 2, 3, 4 atau 5 detik. Berdasarkan informasi dari Percobaan 1, kami mengurangi kontras Weber dari cakram mengembang yang terlihat dalam Percobaan 2 dari -0,22 dari -0,19 hingga mendekati ambang respons kepiting (Gambar A1b ), sementara cakram tak terlihat masih ditampilkan pada kontras Weber sebesar 0,00. Masing-masing dari delapan perlakuan yang digunakan dalam percobaan ini diberi kode alfanumerik, mengikuti sistem yang sama seperti percobaan sebelumnya, tetapi dengan penambahan digit yang merujuk pada offset (dalam detik) saat cakram mulai mengembang (dalam kaitannya dengan awal periode stimulus). Setiap kepiting ( n = 48) diuji sekali dalam percobaan ini dan menerima semua delapan perlakuan dalam satu percobaan. Kami membuat 48 ordo unik dari delapan perlakuan dalam rancangan blok acak seimbang, dengan demikian memastikan bahwa setiap kepiting akan menerima ordo unik dari delapan perlakuan dalam percobaannya.
2.6 Protokol Eksperimen
Setiap percobaan dimulai dengan menambatkan seekor kepiting di atas treadmill dan membiarkannya selama 180 detik untuk menyesuaikan diri dengan latar belakang statis abu-abu dengan kecerahan yang sama dengan latar belakang kaustik. Setelah periode ini, kami mengubah latar belakang ke kondisi kaustik pertama dari periode paparan perlakuan pertama. Setiap perlakuan dimulai dengan periode paparan minimal 90 detik, diikuti oleh periode stimulus. Sepanjang semua percobaan dari Eksperimen 1 dan Eksperimen 2, gambar latar belakang kaustik statis yang disajikan kepada kepiting dipilih secara acak dari 200 bingkai animasi kaustik kami, dengan demikian memastikan variabilitas dan mencegah artefak apa pun yang dapat berasal dari pemilihan satu gambar statis di semua percobaan. Karena penilaian respons kami bergantung pada hewan yang berjalan saat terpapar cakram yang mengembang, kami memperpanjang durasi periode paparan untuk percobaan di mana seekor kepiting diam hingga kepiting mulai berjalan setidaknya selama 5 detik, yang biasanya terjadi dalam < 10 detik. Setelah setiap perlakuan, kepiting terus terpapar pada kondisi kaustik periode stimulus untuk durasi yang singkat (~20 detik; ‘periode persiapan’), sehingga memungkinkan kami untuk menyiapkan pengaturan untuk perlakuan berikutnya dalam skrip MATLAB kami sebelum memulai perlakuan berikutnya. Secara total, setiap perlakuan memakan waktu sekitar 2 menit untuk diselesaikan, sehingga total durasi percobaan kurang dari 20 menit.
2.7 Penilaian Respons
Semua rekaman video diimpor ke MATLAB. Di sini, kami menggunakan skrip khusus untuk mengidentifikasi bunyi bip audio yang dimasukkan ke aliran audio setiap video di awal dan akhir setiap cakram yang mengembang yang disajikan. Dengan menggunakan penanda audio ini, kami memotong rekaman video setiap kepiting menjadi delapan klip video individual, satu untuk setiap perlakuan dalam percobaan. Setiap klip video ini berdurasi 15 detik, yang merekam perilaku kepiting dari 5 detik sebelum hingga 5 detik setelah tampilan cakram yang mengembang (5 detik). Di sudut kanan bawah setiap video ini, kami menambahkan animasi tampilan cakram yang mengembang yang diselaraskan secara temporal, dengan animasi untuk perlakuan cakram yang mengembang yang tampak dan tidak tampak menjadi identik. Dengan demikian, klip video individual ini hanya menunjukkan perilaku kepiting selama presentasi cakram yang mengembang tetapi mengecualikan indikasi apa pun tentang kondisi kaustik dalam periode paparan atau stimulus atau kontras/visibilitas cakram yang mengembang, sehingga memungkinkan kami untuk menilai perilaku respons C. maenas secara buta.
Individu C. maenas yang bergerak menunjukkan respons yang kuat terhadap rangsangan cakram yang mengembang dengan kontras rendah dengan memperlambat atau menghentikan gerakan mereka secara drastis (Drerup dan How 2021 ; Venables et al. 2022 ). Studi kami difokuskan pada apakah kepiting merespons (yaitu, respons absolut) terhadap rangsangan seperti predator yang mendekat berdasarkan paparan kaustik sebelumnya dan saat ini. Oleh karena itu, respons C. maenas terhadap cakram yang mengembang dalam percobaan kami dinilai sebagai data respons biner, dengan kepiting merespons cakram yang mengembang (1; ‘membeku’, ditunjukkan dengan menghentikan (atau memperlambat secara drastis) gerakan mereka) atau tidak merespons cakram yang mengembang (0; ‘berjalan’, ditunjukkan dengan melanjutkan gerakan berjalan mereka). Karena menggunakan rangsangan kontras rendah dalam percobaan kami, kami hanya menilai perilaku berjalan kepiting, sedangkan fitur perilaku lain yang sering terlihat sebagai respons terhadap rangsangan kontras yang lebih tinggi (misalnya, gerakan capit; Drerup dan How 2021 ) tidak dipertimbangkan untuk pendekatan penilaian kami. Respons perilaku hanya disertakan saat terjadi dalam fase ekspansi cakram yang disajikan. Semua video dinilai oleh pengamat yang sama.
2.8 Analisis Statistik
Semua statistik dilakukan dalam R v. 4.2.2 (Tim Inti R 2023 ) dan mencakup model efek campuran linier tergeneralisasi (GLMM) dengan struktur kesalahan keluarga binomial dari paket lme4 (Bates et al. 2015 ). Kami memeriksa asumsi untuk semua GLMM menggunakan paket DHARMa (Hartig 2022 ). Efek signifikan setiap faktor dalam model ditentukan menggunakan panggilan ‘drop1’ dari paket lme4 (Bates et al. 2015 ). Dalam kasus di mana perawatan memiliki efek signifikan pada ukuran respons, kami menggunakan paket emmeans (Lenth 2022 ) bersama dengan kontras yang ditulis khusus untuk menghitung perbedaan berpasangan hanya antara perawatan yang telah ditentukan sebelumnya. Di sini, kami mengoreksi pengujian berganda menggunakan metode Benjamini–Hochberg (Benjamini dan Hochberg 1995 ), yang mengendalikan rasio penemuan palsu menggunakan koreksi Bonferroni yang dimodifikasi secara berurutan. Semua visualisasi dirender menggunakan ggplot2 (Wickham 2016 ).
Untuk Percobaan 1, pertama-tama kami menetapkan apakah perubahan dalam kondisi kaustik dari paparan terhadap periode stimulus menimbulkan respons pada kepiting bahkan tanpa adanya cakram yang terlihat (yaitu, positif palsu). Kami menguji (1) apakah proporsi kepiting yang merespons berbeda antara setiap perlakuan dengan cakram mengembang yang tidak terlihat (kontras Weber 0,00; SSI, MSI, SMI, dan MMI). Untuk melakukannya, kami menjalankan GLMM dengan respons terhadap cakram yang mengembang sebagai variabel respons biner (0: tidak merespons; 1: merespons), perlakuan sebagai efek tetap, dan ID kepiting individu sebagai efek acak. Selanjutnya, kami menghitung perbedaan berpasangan antara keempat perlakuan ini menggunakan kontras yang ditulis khusus. Kami juga menguji (2) apakah proporsi kepiting yang merespons berbeda antara perlakuan dengan cakram mengembang yang terlihat (kontras Weber -0,22; SSV, MSV, SMV, MMV) dan yang tidak terlihat (kontras Weber 0,00; SSI, MSI, SMI, MMI) dengan menjalankan GLMM dengan respons terhadap cakram yang mengembang sebagai variabel respons biner (0: tidak merespons; 1: merespons), visibilitas cakram sebagai efek tetap dua tingkat (terlihat vs. tidak terlihat) dan ID kepiting individu sebagai efek acak. Karena tidak satu pun dari keempat perlakuan dengan cakram yang tidak terlihat menghasilkan proporsi kepiting yang merespons yang berbeda secara statistik (uji 1; Tabel A2 ) dan probabilitas respons keseluruhan dalam perlakuan dengan cakram yang terlihat lebih tinggi daripada dalam perlakuan dengan cakram yang tidak terlihat (uji 2; lihat bagian Hasil), kami menganggap probabilitas respons terhadap perlakuan dengan cakram mengembang yang terlihat tidak terpengaruh oleh respons positif palsu dan dengan demikian menghapus semua perlakuan dengan cakram yang tidak terlihat (SSI, MSI, SMI, dan MMI) dari analisis lebih lanjut. Kami menguji apakah penyertaan atau pengecualian perawatan dengan cakram tak terlihat akan memengaruhi hasil analisis statistik berikutnya secara kualitatif, tetapi menemukan bahwa kedua pendekatan menghasilkan hasil yang konsisten dan serupa secara statistik.
Untuk perawatan yang tersisa yang berisi cakram menjulang yang terlihat (SSV, MSV, SMV dan MMV), kami menguji (3) apakah kemampuan C. maenas untuk merespons dipengaruhi semata-mata oleh keberadaan latar belakang kaustik yang bergerak pada saat penyajian stimulus saja, serta (4) apakah paparan sebelumnya terhadap kaustik yang bergerak memengaruhi kemungkinan respons bahkan setelah gerakan kaustik berhenti. Untuk pengujian 3 dan 4, kami menjalankan GLMM dengan respons terhadap cakram yang mengembang sebagai variabel respons biner (0: tidak merespons; 1: merespons), perawatan sebagai efek tetap dan ID kepiting individu sebagai efek acak. Untuk menentukan (3) apakah semata-mata latar belakang kaustik yang bergerak pada saat penyajian stimulus saja memengaruhi kemampuan C. maenas untuk merespons, kami menghitung kontras berpasangan antara perawatan SSV dan SMV, serta perawatan MSV dan MMV. Dalam kedua perbandingan (SSV vs. SMV; MSV vs. MMV), dua perlakuan yang sesuai hanya berbeda dalam apakah kaustik itu bergerak atau statis ketika stimulus yang menjulang ditunjukkan dalam periode stimulus. Hal ini memungkinkan kami untuk menguji apakah paparan terhadap latar belakang yang bergerak semata-mata selama penyajian cakram yang mengembang memengaruhi kemungkinan merespons stimulus yang menjulang itu. Untuk menetapkan (4) apakah paparan sebelumnya terhadap kaustik yang bergerak saja memengaruhi kemungkinan respons setelah gerakan kaustik berhenti, kami menghitung kontras berpasangan antara perlakuan SSV dan MSV, serta perlakuan SMV dan MMV. Dalam perbandingan ini, dua perlakuan yang sesuai hanya bervariasi dalam kondisi kaustik periode paparan (statis atau bergerak). Oleh karena itu, setiap perbedaan dalam kemungkinan merespons stimulus yang menjulang antara perlakuan ini akan menunjukkan efek historis dari kaustik yang bergerak pada persepsi visual.
Untuk Eksperimen 2, kami juga menetapkan apakah perubahan dalam kondisi kaustik tanpa adanya cakram yang terlihat menimbulkan respons pada kepiting (yaitu, positif palsu). Untuk melakukannya, kami menguji (5) apakah proporsi kepiting yang merespons berbeda antara perlakuan dengan cakram yang terlihat (kontras Weber -0,19; SSV0, MSV0–MSV5) dan yang tidak terlihat (kontras Weber 0,00; MSI0) dengan menjalankan GLMM dengan respons terhadap cakram yang mengembang sebagai variabel respons biner (0: tidak merespons; 1: merespons), visibilitas cakram sebagai efek tetap dua tingkat (terlihat vs. tidak terlihat) dan ID kepiting individu sebagai efek acak. Karena probabilitas respons pada perlakuan dengan cakram yang terlihat lebih tinggi daripada pada perlakuan dengan cakram yang tidak terlihat (lihat bagian Hasil), kami menganggap probabilitas respons terhadap perlakuan dengan cakram yang mengembang tidak terpengaruh oleh respons positif palsu dan dengan demikian menghilangkan perlakuan dengan cakram yang tidak terlihat (MSI0) dari analisis lebih lanjut. Kami menguji apakah penyertaan atau pengecualian perlakuan MSI0 akan memengaruhi hasil analisis statistik berikutnya secara kualitatif, tetapi menemukan bahwa kedua pendekatan menghasilkan hasil yang konsisten dan serupa secara statistik. Untuk perlakuan yang tersisa yang berisi loom yang terlihat (SSV0, MSV0–MSV5), kami kemudian menetapkan (6) berapa lama paparan sebelumnya terhadap kaustik yang bergerak memengaruhi probabilitas respons pada C. maenas . Untuk melakukannya, kami menjalankan GLMM dengan respons terhadap cakram yang mengembang sebagai variabel respons biner (0: tidak merespons; 1: merespons), perlakuan sebagai efek tetap, dan ID kepiting individu sebagai efek acak. Selanjutnya, kami menghitung kontras berpasangan antara SSV0 (paparan kaustik statis berkelanjutan) dan masing-masing dari enam perlakuan MSV (MSV0–MSV5) untuk menentukan kapan tidak ada perbedaan statistik dalam proporsi kepiting yang merespons antara perlakuan.
2.9 Pernyataan Etika
Percobaan yang diuraikan dalam penelitian ini mematuhi pedoman ASAB/ASB untuk penggunaan hewan dalam penelitian perilaku, dilakukan sesuai dengan undang-undang Inggris dan disetujui oleh Badan Peninjauan Kesejahteraan Hewan dan Etika Universitas Bristol (UIN/21/061).
3 Hasil
3.1 Percobaan 1
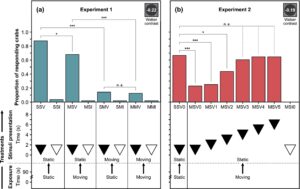
Untuk empat perlakuan dengan cakram yang terlihat (SSV, MSV, SMV dan MMV), paparan terhadap latar belakang kaustik yang bergerak mempengaruhi kemungkinan kepiting untuk merespons rangsangan seperti predator (GLMM: LRT 3 = 108,76; p < 0,001; Gambar 2a ). Lebih sedikit kepiting yang merespons cakram yang mengembang saat terpapar kaustik yang bergerak selama penyajian stimulus, dibandingkan dengan paparan kaustik statis selama penyajian stimulus (SSV vs. SMV: z.ratio = 6,76, p < 0,001; MSV vs. MMV: z.ratio = 5,47, p < 0,001; Gambar 2a ; Tabel A3 ). Ketika terpapar pada kaustik yang bergerak pada saat penyajian cakram, paparan tambahan sebelumnya terhadap kaustik yang bergerak pada periode pengujian tidak mempengaruhi kemungkinan menanggapi rangsangan (SMV vs. MMV, z.ratio = 0,28, p = 0,781; Gambar 2a ; Tabel A3 ). Oleh karena itu, keberadaan gerakan latar belakang kaustik pada saat penyajian isyarat saja mempengaruhi kemampuan kepiting untuk menanggapi rangsangan yang menjulang, terlepas dari paparan kaustik sebelumnya. Selain itu, bahkan ketika ada latar belakang kaustik statis pada saat penyajian cakram, kepiting cenderung tidak menanggapi cakram yang mengembang ini ketika mereka sebelumnya mengalami kaustik yang bergerak (SSV vs. MSV; z.ratio = 2,43; p = 0,02; Gambar 2a ; Tabel A3 ). Oleh karena itu, paparan sebelumnya terhadap gerakan latar belakang kaustik yang bergerak mengurangi kemungkinan kepiting untuk menanggapi rangsangan yang menjulang bahkan setelah kaustik berhenti.
3.2 Percobaan 2
Mengingat bahwa paparan sebelumnya terhadap kaustik yang bergerak mengurangi tingkat respons kepiting terhadap cakram yang mengembang bahkan setelah kaustik berhenti (Eksperimen 1, Gambar 2a ), dalam Eksperimen 2, kami menguji seberapa lama efek ini bertahan. Pergeseran temporal antara akhir paparan kaustik yang bergerak dan awal penyajian cakram memengaruhi kemungkinan C. maenas untuk merespons rangsangan cakram (GLMM: LRT 6 = 54,20; p < 0,001). Paparan sebelumnya terhadap kaustik yang bergerak mengganggu kemampuan kepiting pantai untuk merespons cakram yang mengembang seperti predator yang muncul hingga 2 detik setelah gerakan latar belakang kaustik berhenti (SSV0 vs. MSV0: z = 4,49, p < 0,001; SSV0 vs. MSV1: z = 4,29, p < 0,001; SSV0 vs. MSV2: z = 2,43, p = 0,03; Gambar 2b ; Tabel A4 ). Namun, tidak ada perbedaan dalam proporsi kepiting yang merespons cakram yang mengembang mulai setidaknya 3 detik setelah akhir tampilan kaustik yang bergerak, dibandingkan dengan perawatan kontrol statis SSV0 (Gambar 2b ; Tabel A4 ). Temuan kami menunjukkan bahwa paparan sebelumnya terhadap bahan kaustik dapat mengurangi kemungkinan C. maenas merespons stimulus kontras rendah (kontras Weber -0,19) dalam meniru predator yang mendekat hingga sekitar 65% (32 dari 48 [SSV0] vs. 11 dari 48 kepiting yang merespons [MSV0]; Gambar 2b ) segera setelah bahan kaustik berhenti, dengan efek ini berlangsung setidaknya 2 detik untuk jenis stimulus ini.
4 Diskusi
Gerakan latar belakang kaustik mengurangi tingkat respons C. maenas terhadap rangsangan seperti predator yang mendekat. Sementara temuan kami mengonfirmasi pengamatan terbaru bahwa kaustik yang bergerak secara dinamis dapat mengganggu kemampuan spesies akuatik untuk merespons objek yang bergerak (Attwell et al. 2021 ; Matchette et al. 2020 ; Venables et al. 2022 ), kami juga menetapkan bahwa persepsi visual yang berkurang ini sebagian besar disebabkan oleh gerakan latar belakang pada saat objek disajikan saja, terlepas dari apakah kepiting terpapar kaustik yang bergerak sebelumnya atau tidak. Selain itu, kami menetapkan bahwa paparan sebelumnya terhadap kaustik yang bergerak memiliki efek yang bertahan lama pada kemungkinan merespons objek yang bergerak di C. maenas bahkan setelah paparan kaustik yang bergerak telah berhenti. Secara khusus, kepiting menunjukkan tingkat respons yang berkurang untuk rangsangan yang mendekat yang dimulai hingga 2 detik setelah kaustik yang bergerak berakhir. Oleh karena itu, penelitian kami menunjukkan bahwa tidak hanya kehadiran gerakan latar belakang kaustik dapat mengganggu tugas yang dipandu secara visual pada C. maenas tetapi juga paparan kaustik sebelumnya dapat mengurangi kemungkinan untuk merespons isyarat visual, bahkan setelah paparan kaustik berhenti. Meskipun kaustik dapat merusak tingkat respons objek pada hewan akuatik (studi saat ini; Attwell et al. 2021 ; Matchette et al. 2020 ; Venables et al. 2022 ), sifat-sifat pasti dari kaustik yang menyebabkan penurunan kemampuan visual ini masih belum sepenuhnya dipahami. Pita cahaya kaustik dapat memengaruhi pemandangan visual dengan menciptakan gerakan latar belakang. Sementara gerakan latar belakang dapat menurunkan kemampuan sistem visual untuk memisahkan isyarat spasiotemporal dari objek yang ditargetkan terhadap latar belakangnya (Churan dan Ilg 2002 ), itu juga dapat bertindak sebagai kendala pada proses kognitif dengan mengalihkan atau menyesatkan perhatian pengamat (Dominoni et al. 2020 ). Oleh karena itu, banyak hewan telah mengembangkan mekanisme yang memungkinkan mereka untuk mengurangi dampak gerakan yang mungkin terjadi pada persepsi mereka (Clifford et al. 2007 ; Kohn 2007 ; Webster 2015 ). Misalnya, sistem visual mungkin mengalami berbagai jenis adaptasi gerakan, yang didefinisikan sebagai penurunan sementara aktivitas saraf terhadap stimulus konstan (Barlow dan Hill 1963 ). Dengan beradaptasi dengan stimulus gerakan konstan, sistem visual dapat membatalkan stimulus ini dengan menetapkannya sebagai norma yang diharapkan dari pemandangan visual, dengan demikian meningkatkan kemungkinan mendeteksi stimulus gerakan lain yang berbeda atau baru dengan pemandangan visual yang diadaptasi (misalnya, predator yang mendekat dengan latar belakang yang bergerak) (Webster 2015 ). Mempertimbangkan bahwa kaustik tidak memiliki komponen arah pada skala spasial dan temporal yang luas, dengan pita cahaya bergerak ke segala arah melintasi substrat, C. maenas mungkin melakukan pengurangan perolehan gerakan, mirip dengan pengurangan perolehan kontras yang diamati pada lalat (Harris et al. 2000 ), untuk beradaptasi dengan gerakan latar belakang multiarah yang disediakan oleh kaustik. Adaptasi gerakan ini dapat membantu mencegah saturasi neuron deteksi gerakan, dengan demikian berpotensi memungkinkan kepiting untuk sedikit mengurangi dampak gerakan latar belakang kaustik dengan mencoba meningkatkan kepekaan mereka terhadap bentuk gerakan lainnya. Pengurangan perolehan kontras pada lalat merupakan adaptasi visual yang cepat dan dapat terjadi dalam waktu kurang dari 35 ms setelah terpapar stimulus bergerak (Nordström et al. 2011 ). Jika C. maenas mengalami jenis adaptasi yang sama sebagai respons terhadap gerakan latar belakang yang bersifat kaustik, hal itu kemungkinan terjadi dalam jangka waktu yang sangat singkat (puluhan hingga ratusan milidetik) yang tidak dapat dideteksi oleh pengamatan perilaku kami (dan sebaliknya memerlukan pengukuran intraseluler), yang menjelaskan mengapa kami tidak menemukan perbedaan dalam tingkat respons antara perlakuan SMV dan MMV. Aspek dinamis lain yang dikenakan bahan kaustik pada pemandangan visual hewan adalah fluktuasi intensitas cahaya. Pita cahaya kaustik dapat menghasilkan puncak iluminasi saat bergerak melintasi latar belakang visual (Drerup, Dunkley, et al. 2024 ; Venables et al. 2022 ) atau langsung melintasi mata hewan. Oleh karena itu, krustasea yang terpapar bahan kaustik dapat menyesuaikan sistem visual mereka dengan mengurangi jumlah cahaya yang masuk, misalnya, melalui migrasi butiran pigmen penyaring (Meyer-Rochow 1999 , 2001 ). Namun, mengurangi asupan cahaya dapat menurunkan sensitivitas visual C. maenas (Brodrick et al. 2022 ). Meskipun kepiting yang diuji dalam tatanan eksperimen kami terus-menerus terpapar pada latar belakang kaustik, latar belakang tersebut bervariasi antara diam atau bergerak. Karena latar belakang kaustik yang diam hanya menghasilkan beberapa puncak iluminasi yang terdistribusi secara spasial namun tidak bergerak dan dengan asumsi bahwa kepiting tidak menggerakkan mata mereka sebagai respons terhadap paparan kaustik statis, hanya area retina yang terpapar puncak iluminasi ini dalam bidang visual yang perlu melakukan adaptasi kecerahan. Sebaliknya, untuk latar belakang kaustik yang bergerak, puncak iluminasi bergerak melintasi seluruh bidang visual, sehingga membutuhkan area retina kepiting yang lebih luas untuk beradaptasi dengan peningkatan tingkat kecerahan. Oleh karena itu, kaustik yang bergerak dapat mengurangi sensitivitas visual sebagian besar sistem visual kepiting, yang berpotensi menjelaskan tingkat respons yang berkurang terhadap stimulus mirip predator terhadap latar belakang kaustik yang bergerak. Sementara sistem visual C. maenas berpotensi mengalami adaptasi dalam menanggapi bahan kaustik yang bergerak, contoh dari penglihatan manusia telah menunjukkan bahwa adaptasi visual juga dapat memengaruhi sistem visual bahkan setelah paparan adaptor berakhir (Thompson dan Burr 2009 ) hingga 14 detik (Ashida dan Osaka 1994 ). Contoh paling terkenal dari efek samping tersebut adalah ‘ilusi air terjun’ (Addams 1834 ), yang menggambarkan bahwa setelah paparan yang lama pada pemandangan visual yang bergerak dalam arah tertentu (misalnya, air terjun), objek yang diam dapat tampak bergerak ke arah yang berlawanan (Anstis et al. 1998 ). Mempertimbangkan bahwa setelah terpapar bahan kaustik yang bergerak, kemungkinan respons C. maenas untuk mendeteksi rangsangan seperti predator berkurang hingga 2 detik, dapat dibayangkan bahwa pengurangan sementara dalam kemampuan untuk merespons objek ini disebabkan oleh efek samping yang disebabkan oleh adaptasi visual dalam menanggapi paparan bahan kaustik sebelumnya. Efek samping gerakan yang dijelaskan sebelumnya, seperti ilusi air terjun, merupakan hasil dari paparan terhadap rangsangan visual yang bergerak dalam satu arah (Thompson dan Burr 2009 ). Oleh karena itu, bahan kaustik kemungkinan tidak dapat menimbulkan respons saraf yang sama karena gerakan multiarahnya. Namun, jika C. maenas melakukan pengurangan perolehan gerakan sebagai respons terhadap gerakan bahan kaustik, adaptasi visual ini berpotensi menghasilkan efek samping visual historis, yang dapat menyebabkan berkurangnya tingkat respons terhadap rangsangan mirip predator pada spesies ini. Selain itu, dapat dibayangkan juga bahwa adaptasi kecerahan potensial terhadap bahan kaustik yang bergerak dapat menimbulkan efek samping yang mengakibatkan berkurangnya tingkat respons terhadap rangsangan cakram kita. Oleh karena itu, penelitian di masa mendatang harus menetapkan sifat visual mana dari paparan bahan kaustik yang memengaruhi sistem visual hewan selama dan setelah paparan bahan kaustik. Sementara dasar neural dari efek samping visual dan manfaat serta konsekuensi teoritisnya pada sistem visual baru-baru ini telah diselidiki (Glasser et al. 2011 ; Webster 2015 ), pekerjaan sebelumnya sering kali berfokus pada rangsangan gerakan yang terstandarisasi namun tidak realistis secara ekologis yang tidak akan dialami hewan di lingkungan alaminya, seperti kisi-kisi gelombang sinus yang melayang (misalnya, Glasser et al. 2011 ; Kohn dan Movshon 2003 ). Sebaliknya, penelitian kami menunjukkan bahwa gerakan latar belakang yang disebabkan oleh pola pencahayaan kaustik, bentuk umum dari iluminasi dinamis di habitat laut dangkal, dapat menyebabkan efek samping visual pada kepiting pantai yang dapat mengurangi deteksi objek mereka. Secara khusus, pengaturan eksperimen kami meniru skenario di mana seekor hewan didekati oleh predator dengan latar belakang vertikal atau miring, seperti yang dapat ditemukan di habitat pesisir dangkal atau kolam batu besar. Apakah efek historis paparan bahan kimia kaustik ini memiliki konsekuensi terhadap risiko pemangsaan C. maenas , atau hanya merupakan produk sampingan dari adaptasi visual, masih belum diketahui. Secara khusus, memanfaatkan efek historis yang mungkin dimiliki bahan kimia kaustik pada sistem visual mangsa mungkin hanya dapat dilakukan oleh predator yang tidak terpengaruh oleh bahan kimia kaustik, seperti predator burung yang terbang yang persepsi visualnya kemungkinan tidak terhalang oleh bahan kimia kaustik saat mencari mangsa dari atas permukaan air. Predator ini dapat mengatur waktu serangan mencari makan untuk menyerang mangsa, seperti C. maenas, dalam beberapa saat setelah bahan kimia kaustik berhenti mendapatkan manfaat dari berkurangnya tingkat deteksi objek pada mangsanya. Penelitian di masa mendatang harus menetapkan apakah predator di alam liar benar-benar dapat meningkatkan keberhasilan berburu mereka dengan menargetkan mangsanya dalam beberapa saat setelah bahan kimia kaustik berakhir. Sebagai alternatif , versi adaptasi dari pengaturan eksperimen kami dapat digunakan untuk mengekspos predator nyata pada paparan bahan kimia kaustik dan rangsangan mangsa yang dihasilkan komputer untuk menguji apakah predator ini mengatur waktu serangan mencari makan mereka pada periode singkat setelah bahan kimia kaustik berakhir. Temuan penelitian ini membuka jalan penelitian yang menarik untuk menyelidiki lebih lanjut bagaimana paparan kaustik dan gerakan latar belakang secara umum memengaruhi sistem visual hewan. Misalnya, kepiting dalam pengaturan eksperimen kami terpapar durasi tetap > 90 detik gerakan latar belakang kaustik, sedangkan di habitat alami, periode paparan kaustik dapat bervariasi panjangnya dari beberapa detik hingga beberapa jam. Oleh karena itu, penelitian di masa mendatang dapat mengeksplorasi efek dari waktu paparan yang lebih pendek dan lebih lama pada efek historis kaustik, misalnya, untuk menetapkan waktu paparan minimum yang diperlukan untuk memperoleh adaptasi visual ini atau untuk menentukan apakah periode paparan yang lebih lama memperpanjang durasi efek historis ini. Arah penelitian potensial lainnya dapat mengeksplorasi apakah, dan sejauh mana, tingkat intensitas dan kecepatan gerakan kaustik memengaruhi keberadaan dan durasi efek historis dalam sistem visual hewan, dan bagaimana parameter tersebut berinteraksi dengan perubahan analog dalam stimulus cakram yang disajikan (misalnya, perbedaan dalam kekuatan kontras dan kecepatan perluasan stimulus seperti predator). Misalnya, jika sistem visual seekor kepiting yang terpapar pada latar belakang kaustik yang bergerak beradaptasi dengan kecepatan gerak rata-rata yang dirasakan dalam pemandangan visual, efek historis yang diamati dalam bentuk tingkat respons objek yang berkurang mungkin lebih lemah atau berlangsung dalam durasi yang lebih pendek jika kecepatan objek yang mendekat menyimpang secara signifikan dari kecepatan yang disesuaikan.
Singkatnya, temuan kami menunjukkan bahwa kaustik bergerak yang hanya disajikan pada presentasi stimulus dapat mengurangi kemampuan C. maenas untuk merespons stimulus ini; namun, paparan kaustik sebelumnya sebagai tambahan terhadap paparan kaustik ini tidak meningkatkan atau menurunkan tingkat respons terhadap stimulus seperti predator pada spesies ini. Selain itu, kami menemukan bahwa paparan kaustik sebelumnya dapat memiliki efek historis jangka pendek pada persepsi visual di C. maenas , mengurangi tingkat respons visualnya secara signifikan setidaknya selama dua detik setelah kaustik berakhir. Studi kami memotivasi pekerjaan masa depan tentang bagaimana gerakan latar belakang yang terjadi secara alami memengaruhi sistem visual hewan dan adaptasi sensorik atau perilaku mana yang telah berevolusi untuk mengurangi atau memanfaatkan efek historis pada persepsi visual.