
ABSTRAK
Tujuan
Perubahan iklim merupakan salah satu ancaman utama bagi keanekaragaman hayati global, dan perubahan tersebut diperkirakan akan menyebabkan pergeseran rentang distribusi dan pola keanekaragaman. Kami mengevaluasi apakah kawasan lindung dan sisa hutan di Hutan Atlantik di Amerika Selatan (AF) diproyeksikan dapat memastikan keanekaragaman taksonomi (TD) dan keanekaragaman filogenetik (PD) mamalia kecil yang tidak hidup di bawah skenario perubahan iklim di masa mendatang.
Lokasi
Hutan Atlantik (AF), Amerika Selatan.
Metode
Kami menggunakan Species Distribution Modelling (SDMs) melalui pendekatan ensemble untuk menilai potensi distribusi 101 spesies mamalia kecil menggunakan skenario iklim saat ini (1979–2013) dan masa depan (2050 dan 2070). Kami mempertimbangkan skenario konsentrasi gas rumah kaca yang optimis dan pesimis (SSP370 dan SSP585). Kami mengakses TD melalui jumlah area yang sesuai vs. area dengan peta distribusi kesesuaian yang rendah atau tidak diketahui untuk setiap spesies dan PD menggunakan jumlah panjang cabang pohon filogenetik yang dispasialkan.
Hasil
Model kami menunjukkan bahwa perubahan iklim kemungkinan akan mengurangi daerah beriklim cocok untuk mamalia kecil di AF. Penyusutan dalam distribusi potensial diproyeksikan akan menyebabkan hilangnya TD dan PD yang tinggi. Wilayah tenggara Hutan Atlantik kemungkinan akan mengalami penurunan PD yang paling nyata, sementara beberapa daerah kecil di Hutan Atlantik selatan diproyeksikan akan meningkatkan PD di masa mendatang.
Kesimpulan Utama
Model kami menunjukkan penurunan tajam dalam TD dan PD dari daerah hotspot keanekaragaman hayati di AF dalam skenario perubahan iklim. Karena mamalia kecil memiliki kemampuan penyebaran yang rendah, dan karena sebagian besar AF sangat terfragmentasi, kecil kemungkinan bioma ini akan mempertahankan keanekaragaman hayati mamalia kecil di masa mendatang.
1 Pendahuluan
Perubahan iklim diperkirakan akan mendorong pergeseran jangkauan spesies dan mengubah pola keanekaragaman dan distribusi (Mota et al. 2022 ; Alves-Ferreira, Giné, et al. 2022 ; Biber et al. 2023 ). Diperkirakan hingga 49% serangga, 44% tumbuhan, dan 26% vertebrata akan kehilangan lebih dari setengah jangkauan geografis historisnya (Wudu et al. 2023 ), yang berpotensi menyebabkan penurunan kekayaan spesies di masa depan dan pemiskinan komunitas. Sementara jumlah spesies (Keanekaragaman Taksonomi; TD) banyak digunakan untuk mengevaluasi dampak perubahan iklim terhadap keanekaragaman hayati (misalnya, Mota et al. 2022 ), hal itu tidak memperhitungkan sejarah evolusi di antara spesies. Hal ini menyoroti pentingnya mempertimbangkan berbagai aspek keanekaragaman, terutama ketika aspek-aspek ini berbeda (Mendes et al. 2020 ).
Respons terhadap perubahan iklim berbeda di antara spesies, dan beberapa mungkin lebih rentan daripada yang lain, yang menunjukkan bahwa hilangnya spesies karena penyebab iklim tidak didistribusikan secara acak di seluruh filogeni (Prinzing et al. 2001 ; Thuiller et al. 2005 ). Faktanya, respons distribusi spesies terhadap perubahan iklim cenderung lebih mirip antara spesies yang terkait erat (Eiserhardt et al. 2015 ). Akibatnya, kontraksi rentang distribusi di antara spesies-spesies ini dapat mengakibatkan hilangnya klade yang berbeda dan tak tergantikan, sehingga mengurangi keanekaragaman filogenetik (PD) (Thuiller et al. 2011 ). Misalnya, proyeksi masa depan menunjukkan pengurangan PD sebesar 2,7%, 11,5%, dan 9,6% untuk tumbuhan, burung, dan mamalia, masing-masing, dan prevalensi hilangnya spesies berpotensi menyebabkan homogenisasi komunitas Eropa (Thuiller et al. 2011 ). Lebih jauh lagi, tempat-tempat dengan keragaman garis keturunan kuno yang tinggi diproyeksikan akan menghilang atau berpindah lokasi karena menyusutnya jangkauan geografisnya, sehingga mengancam seluruh klade pada pohon filogenetik (González-Orozco et al. 2016 ).
Khususnya, Hutan Atlantik (selanjutnya AF) adalah hutan hujan utama kedua di Amerika, setelah domain Amazon, yang terbentuk antara 60 dan 65 juta tahun yang lalu [mya] (Pennington et al. 2006 ; Colombo dan Joly 2010 ), tetapi berkurang (23% hingga 36%) dan sangat terfragmentasi (~97% fragmen kurang dari 50 ha) (Vancine et al. 2024 ). AF menampung salah satu keanekaragaman fauna terpenting di dunia (Myers et al. 2000 ) dan masih menderita degradasi yang mengancam keanekaragaman hayati. Pemeliharaan kawasan lindung saat ini dan penciptaan yang baru sedang diupayakan oleh banyak peneliti sebagai alternatif untuk melestarikan spesies yang terancam punah dan mengurangi penurunan keanekaragaman hayati (misalnya, Vale et al. 2018 ; Mota et al. 2018 ). Bahasa Indonesia: Terlepas dari hasil positif yang dihasilkan dari kawasan lindung, terutama dalam mencegah penggundulan hutan, kawasan tersebut tetap kurang terwakili (Geldmann et al. 2013 ; Françoso et al. 2015 ). Misalnya, penilaian terkini kawasan endemisme (yaitu, kawasan dengan jumlah spesies unik yang ditemukan tinggi) dalam AF untuk mamalia kecil yang tidak hidup liar telah menunjukkan bahwa sebagian besar kawasan ini tidak dilindungi (Dalapicolla et al. 2021 ). Lebih jauh lagi, kawasan dengan nilai konservasi tinggi yang memiliki keanekaragaman taksonomi dan filogenetik pohon angiospermae yang berbeda di AF juga perlu mendapat perhatian (Saraiva et al. 2018 ). Kawasan yang tidak dilindungi yang telah diabaikan memiliki potensi besar karena kawasan ini mungkin menampung spesies baru yang sudah terancam (Avigliano et al. 2019 ). Sementara beberapa kawasan lindung mungkin tidak lagi cocok untuk spesies tertentu karena perubahan iklim, ada kawasan lindung lain yang diperkirakan akan mempertahankan kondisi iklim yang sama (Ferro et al. 2014 ). Selain itu, kawasan stabil yang tidak dilindungi dapat dipertimbangkan untuk dilindungi di masa depan ketika ada kesempatan (Vale et al. 2018 ), untuk AF (Tonetti et al. 2024 ).
Satu ukuran keanekaragaman yang menjadi populer pada awal abad ke-21 adalah keanekaragaman filogenetik (selanjutnya disebut sebagai PD; Faith 1992 ), yang digunakan secara luas menggunakan informasi genetik untuk memulihkan hubungan filetik. PD menyampaikan informasi mengenai proses evolusi yang mendorong struktur komunitas (Dreiss et al. 2015 ; Safi et al. 2011 ). Ini khususnya berguna untuk spesies dengan relung ekologi yang serupa (Kozak dan Wiens 2006 ; Wiens dan Graham 2005 ), karena struktur filogenetik komunitas dapat menawarkan wawasan ke dalam persistensi evolusi spesies dan proses biogeografi yang membentuk komposisi komunitas. Mengenali pola distribusi spesies adalah langkah pertama untuk melakukan analisis biogeografi apa pun karena pola-pola ini memberikan informasi dasar untuk menguji hipotesis proses biogeografi historis atau ekologis (Myers dan Giller 1988 ).
Model Distribusi Spesies (SDM) telah semakin banyak digunakan untuk mengevaluasi dampak perubahan iklim terhadap distribusi spesies (Loyola et al. 2014 ; Zwiener et al. 2018 ; de Lima et al. 2019 ), dan untuk memandu langkah-langkah konservasi (Vale et al. 2018 ). Pendekatan pemodelan telah menunjukkan peningkatan berkelanjutan, dengan praktik dan pedoman yang baik tersedia (misalnya, Zurell et al. 2020 ; Sillero et al. 2021 ), serta metodologi yang sangat kuat untuk memprediksi distribusi spesies dan perubahan komposisi komunitas (misalnya, Alves-Ferreira, Giné, et al. 2022 ; Alves-Ferreira, Talora, et al. 2022 ; Mota et al. 2022 ; de Oliveira-Silva et al. 2022 ). SDM telah terbukti menjadi alat yang berguna dalam mengidentifikasi area-area penting untuk perlindungan keanekaragaman spesies (Bonfim et al. 2019 ). Meskipun terdapat area-area yang secara historis stabil yang terletak di AF, selain tingkat PD yang tinggi (Pugliesi dan Rapini 2015 ; de Oliveira Bünger et al. 2016 ), wilayah tersebut masih kekurangan informasi tentang bagaimana pohon kehidupan akan berubah di masa depan. Memahami apakah area-area yang relevan untuk konservasi di dalam salah satu hutan paling terancam di planet ini akan berfungsi sebagai tempat perlindungan bagi sejarah evolusi di masa depan adalah hal yang sangat penting (Vale et al. 2021 ).
Di sini, kami fokus pada mamalia kecil (tikus dan marsupial) dari AF untuk menilai apakah sisa-sisa hutan dan kawasan lindung dapat mempertahankan keanekaragaman taksonomi (TD) dan keanekaragaman filogenetik (PD) dalam menghadapi perubahan iklim. Mengingat kontraksi habitat yang sesuai secara iklim yang diharapkan untuk spesies ini, kami berhipotesis penurunan yang signifikan dalam TD dan PD di seluruh wilayah. Mamalia kecil dipilih sebagai kelompok fokus karena kapasitas penyebarannya yang terbatas (Bowman et al. 2002 ), yang dapat menghambat kemampuan mereka untuk melacak perubahan kondisi iklim dan menjajah habitat yang baru cocok (Fricke et al. 2022 ). Dengan demikian, studi ini meningkatkan pemahaman kita tentang dampak perubahan iklim pada TD dan PD mamalia kecil, memberikan wawasan baru tentang potensi ketahanan kelompok ini dalam lanskap yang terfragmentasi dan peran sisa-sisa hutan dan kawasan lindung dalam melestarikan keanekaragaman hayati di masa depan.
2 Metode
2.1 Lokasi Penelitian
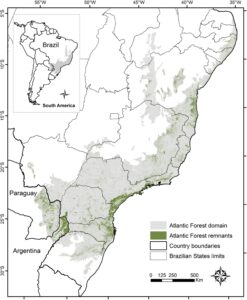
AF dicirikan oleh topografi yang kompleks dan distribusi lintang yang luas di sepanjang pantai Brasil (dari 3° hingga 33°) dan pedalaman Argentina dan Paraguay (Gambar 1 ) (de Muylaert et al. 2018 ). Rentang lintang yang luas dikombinasikan dengan gradien ketinggian menghasilkan berbagai macam formasi floristik, termasuk hutan hujan dataran rendah dan pegunungan yang selalu hijau, hutan gugur dan semi-gugur, hutan Araucaria subtropis, dan hutan ‘brejos de altitudes’ (Oliveira-Filho dan Fontes 2000 ; da Silva et al. 2004 ; Joly et al. 2014 ). Hutan-hutan ini menyediakan layanan ekosistem penting (misalnya, penyerapan karbon, pemurnian air dan pemeliharaan kesuburan tanah) untuk salah satu daerah terpadat di Brasil (Ditt et al. 2010 ). Hutan dulunya menutupi lebih dari 150 juta hektare tetapi saat ini hanya ~23% dari area hutan asli yang tersisa dalam lanskap terfragmentasi yang didominasi oleh area antropogenik (Vancine et al. 2024 ). Saat ini, sisa-sisa hutan ini terancam oleh perburuan liar (Galetti et al. 2009 , 2017 ), penebangan (Chiarello 1999 ), dan infrastruktur buatan manusia (misalnya, jaringan pipa, jalan), yang semuanya meningkatkan terjadinya kematian mamalia besar (Miotto et al. 2012 ; Galetti et al. 2017 ). Di wilayah ini, kami fokus pada mamalia kecil terestrial yaitu sekitar 30 spesies marsupial (Didelphidae) dan 97 spesies hewan pengerat kecil (Caviidae, Cricetidae, Ctenomyidae, Echimyidae) (Bovendorp et al. 2017 ; Dalapicolla et al. 2021 ; Abreu et al. 2022 ).
2.2 Catatan Kejadian dan Variabel Bioklimat
Kami mengumpulkan catatan kejadian dari literatur (Dalapicolla et al. 2021 ) dan dari platform online: GBIF (www.gbif.org , 2024), spesiesLink (www.spesieslink.net , 2024), VertNet ( https://www.vertnet.org ) , iDigBio ( https://www.idigbio.org ), Sistema da Informação sobre a Biodiversidade Brasileira (SiBBr) ( www.sibbr.gov.br , 2024), iDigBio ( www.idigbio.org/ , 2024), dan Portal da Biodiversidade–ICMBio ( https://portaldabiodiversidade.icmbio.gov.br/portal/ , 2024). Pencarian menghasilkan 129.841 kemunculan untuk 107 spesies. Setelah itu, kami melakukan kontrol kualitas pada kejadian untuk menghilangkan duplikat, rekaman yang tidak mungkin atau tidak mungkin, georeferensi dalam centroid negara, negara bagian, dan kota, atau georeferensi dalam institusi (yaitu, universitas, kebun binatang), hanya menyaring kejadian dalam batas spasial Amerika Selatan, dan menemukan lokasi tersebut di atas lautan melalui paket R CoordinateCleaner (Zizka et al. 2019 ). Kami menggunakan rekaman kejadian yang dikumpulkan antara tahun 1970 dan 2024. Masalah taksonomi dalam nama spesies dikoreksi oleh spesialis mamalia kecil. Kami menipiskan kejadian untuk mengurangi bias spasial menggunakan penyaringan 5 km melalui paket R spThin (Aiello-Lammens et al. 2015 ). Kami juga menerapkan filter lingkungan dengan variabel bioklimatik untuk menghilangkan rekaman kejadian dengan nilai yang sama untuk kondisi lingkungan yang sama (menggunakan 12 kelas) dengan paket R flexsdm (Velazco et al. 2022 ). Kemudian, kami memilih spesies dengan lebih dari 10 rekaman kejadian untuk melakukan analisis lebih lanjut. Basis data terakhir kami terdiri dari 12.166 catatan kejadian untuk 101 spesies mamalia kecil (Tabel S1 ; Gambar S1 ).
Kami memperoleh 19 variabel bioklimat (BIO01 hingga BIO19) dari basis data CHELSA v.2.1 (Karger et al. 2017 ) dengan resolusi 2,5 arc-min (~5 km 2 ) untuk wilayah spasial Amerika Selatan. Kami menggunakan skenario iklim saat ini (1979–2013) dan dua skenario iklim masa depan: 2050 dan 2070. Untuk setiap skenario masa depan, kami mempertimbangkan dua Shared Socioeconomic Pathways (SSP) yang mewakili skenario gas rumah kaca optimis (SSP370) dan pesimis (SSP585). Kami menggunakan semua Model Sirkulasi Global (GCM) yang tersedia di CHELSA: GFDL-ESM4, MPI-ESM1-2-HR, MRI-ESM2-0, IPSL-CM6A-LR, dan UKESM1-0-LL (Navarro-Racines et al. 2020 ; Cannon 2020 ) untuk meminimalkan ketidakpastian tentang pilihan hanya satu GCM (Thuiller et al. 2019 ) dan membuat raster rata-rata dengan lima GCM untuk dua Jalur Sosial Ekonomi Bersama (SSP).
2.3 Pemodelan Distribusi Spesies
Kami menggunakan Model Distribusi Spesies (SDM) untuk memodelkan area mamalia kecil yang sesuai secara iklim dengan mengaitkan kejadian dan serangkaian variabel bioklimat. Kami menjelaskan metodologi untuk SDM mengikuti protokol standar ODMAP (Gambaran Umum, Data, Model, Penilaian, dan Prediksi) (Zurell et al. 2020 ). Di sini, kami menyertakan ringkasan bagian ‘gambaran umum’, tetapi informasi terperinci tentang setiap langkah pemodelan dapat ditemukan di Informasi Pendukung S2 .
Dalam studi ini, tujuan kami adalah untuk memproyeksikan pola TD dan PD dari 101 spesies mamalia kecil yang tidak terbang untuk skenario perubahan iklim saat ini dan di masa mendatang. Untuk melakukannya, kami melanjutkan sebagai berikut: (1) kami memperoleh kejadian untuk mamalia kecil dan serangkaian prediktor bioklimatik; (2) kami memperoleh beberapa SDM untuk setiap spesies yang dihasilkan dengan beberapa algoritma dan disetel dengan hiperparameter; (3) kami memperoleh model ansambel untuk setiap spesies; (4) kami menumpuk area yang sesuai vs. area peta kesesuaian rendah atau tidak diketahui dan menghitung taksonomi dan PD untuk setiap piksel. Area studi utama kami adalah AF di Amerika Selatan. Namun, kami melakukan kalibrasi dan proyeksi SDM dengan mempertimbangkan seluruh wilayah Amerika Selatan (Garis Bujur -109.446 hingga -26.241, Garis Lintang -58.498 hingga 12.590), karena banyak spesies yang kami evaluasi tidak hanya terdapat di AF, tetapi juga di bioma lain di Amerika Selatan. Batas komunitas ditetapkan sebagai sel grid dengan resolusi 2,5 arc-min (~5 km 2 ). Kami menyesuaikan variabel bioklimat untuk setiap area kalibrasi spesies menggunakan buffer dengan radius ~300 km di sekitar kemunculan spesies (Barve et al. 2011 ; Whitford et al. 2024 ) dan memilih serangkaian variabel dengan VIF lebih rendah dari 2,0 (Dormann et al. 2013 ). Variabel yang sama digunakan untuk memproyeksikan estimasi relung untuk skenario perubahan iklim di masa mendatang.
Kami menggunakan SDM untuk spesies dengan lebih dari 20 kejadian (82 spesies, 81%) dan Ensemble of Small Models (ESM) untuk spesies dengan kurang dari 20 dan lebih dari 10 kejadian (19 spesies, 18%) (Breiner et al. 2015 ). Untuk area kalibrasi setiap spesies, kami mengambil sampel titik latar belakang secara acak (jumlah kehadiran yang sama) untuk algoritma Generalised Linear Models (GLM), Generalised Additive Models (GAM), Random Forest (RF), Support Vector Machines (SVM), Neural Networks Models (NNM), dan Generalised Boosted Regression Models (GBM), dan mengambil sampel titik latar belakang secara acak (jumlah kehadiran dikalikan 10; Whitford et al. 2024 ) untuk Gaussian Process Models (GPM) dan algoritma MaxEnt di seluruh area yang digunakan untuk pemasangan model, karena algoritma ini memerlukan informasi lebih lanjut tentang kondisi lingkungan. Model-model tersebut dipasang dan dievaluasi menggunakan paket R flexsdm (Velazco et al. 2022 ) dengan algoritma yang disebutkan sebelumnya, dipasang tanpa penyetelan: Model Linear Umum (GLM, keluarga distribusi dan poli = 2), Model Aditif Umum (GAM, keluarga distribusi binomial dan spline regresi pelat tipis sebagai dasar penghalusan), dan Model Proses Gaussian (GPM); dan dengan penyetelan hiperparameter: Random Forest (RF; menggunakan mtry = (1, 2, 3, 4, 5, 6, 7), yaitu, jumlah variabel yang akan diambil sampelnya secara acak sebagai kandidat pada setiap pemisahan), Support Vector Machines (SVM; menggunakan C = (2, 4, 8, 16, 20), dan sigma = (0,01, 0,1, 0,2, 0,3, 0,4)), MaxEnt (menggunakan pengali regularisasi = (0,5, 1, 1,5, 2, 2,5, 3, 3,5, 4), dan kelas fitur = (l, lq, h, lqh, lqhp, lqhpt)), Neural Networks Models (NNM; menggunakan ukuran = (2, 4, 6, 8, 10), dan jarak peluruhan = (0,001, 0,05, 0,1, 1, 3, 4, 5, dan 10)), dan Generalised Boosted Regression Models (GBM; menggunakan pohon = (20, 50, 100), penyusutan = (0,1, 0,5, 1), dan n.minobsinnode = (1, 3, 5, 7, 9), yaitu, jumlah minimum pengamatan di simpul terminal pohon). Kami membuat SDM dengan penyetelan hiperparameter karena nilai hiperparameter default sering kali tidak mengembalikan model terbaik (Fourcade 2021 ; Morales et al. 2017 ; Vignali et al. 2020 ).
Kami mengevaluasi dan mengkalibrasi model menggunakan validasi silang blok spasial dengan empat partisi (Santini et al. 2021 ). Validasi blok spasial dianggap sebagai pendekatan yang lebih kuat untuk mengevaluasi transferabilitas model (Roberts et al. 2017 ). Kami mempartisi data menggunakan data kehadiran dan latar belakang menggunakan Validasi silang blok spasial, yang digunakan untuk mempartisi data latar belakang. Pertama, kami mencoba mempartisi ke k = 4 bila memungkinkan; jika fungsi mengembalikan kesalahan, kami menguranginya dan mencoba dengan k = 3, dan jika kesalahan tetap ada, kami melakukannya dengan k = 2. Untuk ESM, kami menggunakan partisi acak, dengan k = 3 dan 5 replikasi karena beberapa titik kejadian. Kami mengakses kinerja model dengan menghitung Indeks Boyce. Kami juga melaporkan metrik evaluasi lain untuk memfasilitasi perbandingan dalam literatur, seperti Area Under Curve (AUC), Omission Rate (OR), FPB, dan Indeks Sorensen.
Kami mengatasi ketidakpastian algoritmik dengan menggunakan metode ensemble yang merata-ratakan hasil dari berbagai algoritma (Araújo dan New 2007 ). Model ensemble dihitung sebagai rata-rata tertimbang dari kesesuaian iklim di semua algoritma, menggunakan nilai Boyce sebagai bobot. Kami memproyeksikan model untuk saat ini, 2050, dan 2070 dalam skenario optimis dan pesimis. Kami mengubah model kontinu akhir (saat ini dan masa depan) menjadi peta area yang sesuai vs. area dengan kesesuaian rendah atau tidak diketahui (area yang sesuai = 1 dan area dengan kesesuaian rendah atau tidak diketahui = 0) menggunakan ambang batas yang memaksimalkan jumlah sensitivitas dan spesifisitas (Max SSS) (Liu et al. 2013 , tetapi lihat Liu et al. 2016 untuk batasan metode tersebut). Kami memilih pendekatan ini karena minat kami dalam artikel ini adalah menganalisis komposisi komunitas ekologi potensial (Stacked Species Distribution Modelling—S-SDM; Ferrier dan Guisan 2006 ; Dubuis et al. 2011 ), dan kemudian menghitung TD dan PD untuk setiap komunitas potensial.
Terakhir, kami menerapkan batasan spasial berdasarkan metode posteriori untuk mengurangi prediksi berlebih di area yang sesuai vs. area dengan peta kesesuaian rendah atau tidak diketahui. Kami menggunakan metode “pres” (hanya pembatasan berdasarkan kejadian) untuk menghilangkan prediksi berlebih, yang hanya mempertahankan piksel-piksel tersebut di petak kesesuaian yang mencegat rekaman kejadian yang diketahui, untuk skenario saat ini dan masa mendatang (Mendes et al. 2020 ). Semua analisis yang terkait dengan pemodelan dilakukan dalam paket R flexsdm (Velazco et al. 2022 ).
2.4 Analisis Filogenetik
Untuk analisis filogenetik, kami menghasilkan filogeni molekuler melalui penelitian dan penyusunan sekuens Sitokrom B untuk spesies yang ada di Pusat Informasi Bioteknologi Nasional (NCBI; Bethesda, Maryland, AS). Kami mengecualikan 30 spesies dari analisis yang tidak memiliki sekuens Sitokrom B. Untuk melakukan penyelarasan sekuens untuk spesies, kami menggunakan Geneious (Biomatters Limited) (Kearse et al. 2012 ). Untuk menghasilkan pohon mamalia kecil AF, kami mengimpor penyelarasan spesies ke perangkat lunak MEGA dan menghasilkan jarak berpasangan untuk setiap spesies. Kami membangun pohon filogenetik berdasarkan metode Nei-Gojobori, yang banyak digunakan untuk merekonstruksi filogeni mamalia (Tobe et al. 2010 ). Dengan demikian, pohon filogenetik kami untuk mamalia kecil AF memiliki tingkat kesesuaian yang kuat dengan filogeni lain (Voss dan Jansa 2009 ; Fabre et al. 2012 ; Jansa et al. 2014 ), dan kami menggunakan validasi ahli untuk memastikan pohon filogenetik kami memiliki inferensi taksonomi yang paling hemat (Percequillo et al. 2011 ; Gambar S2 ).
2.5 Keanekaragaman Taksonomi dan Filogenetik
Kami memperkirakan TD sebagai jumlah spesies di setiap sel grid menggunakan paket R divraster (Mota et al. 2023 ), dan menghitung PD berdasarkan jumlah panjang cabang (Faith 1992 ) untuk spesies di setiap sel grid menggunakan paket R phyloraster (Alves-Ferreira et al. 2024 ). TD dan PD dihitung untuk saat ini, 2050 SSP370, 2050 SSP585, 2070 SSP370, dan 2070 SSP585. Kami tumpang tindih raster TD dan PD dengan kawasan lindung dari World Database on Protected Areas (WDPA 2024 ) dan sisa hutan untuk AF (ATLANTIC SPATIAL, Vancine et al. 2023 ).
Karena PD dapat sangat berkorelasi dengan TD, kami menghitung ukuran efek terstandarisasi (SES) dari PD menggunakan paket R SESraster (Heming et al. 2023 ) dan phyloraster (Alves-Ferreira et al. 2024 ). Untuk ini, kami membandingkan nilai PD yang diamati dengan nilai acak metrik yang dihasilkan dengan 999 pengacakan. Untuk menghasilkan komunitas acak, kami menggunakan algoritma yang mempertahankan kekayaan spesies untuk setiap sel grid dan jumlah situs yang ditempati oleh setiap spesies, sambil mengacak lokasi distribusi setiap spesies (Strona et al. 2014 ). Kemudian, kami menghitung SES menggunakan rumus berikut:
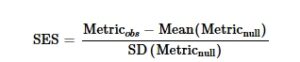
di mana Metric obs adalah nilai yang diamati untuk metrik, mean(Metric rand ) adalah mean metrik yang dihitung berdasarkan 999 randomisasi, dan SD(Metric rand ) adalah simpangan baku dari 999 randomisasi. Nilai positif SES menunjukkan wilayah di mana PD yang diamati lebih tinggi dari yang diharapkan, dan nilai negatif SES menunjukkan wilayah di mana nilai yang diamati lebih rendah dari yang diharapkan secara acak.
3 Hasil
Model ensemble menunjukkan kinerja prediktif sedang hingga tinggi (median Indeks Boyce = 0,910, SD = 0,067; median Indeks Sorensen = 0,667, SD = 0,123; median AUC = 0,698, SD = 0,095; median TSS = 0,434, SD = 0,138; Median FPB = 1,067, SD = 0,259) (lihat Tabel S2 ). Model-model tersebut memproyeksikan penurunan area yang sesuai untuk banyak spesies (59 spesies, 57,84%) sebagai respons terhadap perubahan iklim di masa mendatang. Tiga spesies yang diproyeksikan mengalami penurunan tertinggi dalam distribusi potensial adalah Nectomys squamipes , Caluromys philander , dan Thylamys velutinus , sementara spesies yang diproyeksikan mengalami peningkatan tertinggi dalam kesesuaian adalah Calomys expulsus , Caluromys lanatus , dan Rhipidomys macrurus (Tabel S3 ; Gambar S3 ).
Studi kami memperkirakan bahwa banyak wilayah yang terletak di dalam pusat endemisme mamalia kecil di AF (Gambar S4 ) diproyeksikan akan kehilangan TD di masa mendatang (Gambar 2b–e ). Saat ini, model memproyeksikan TD tinggi di wilayah tenggara AF, termasuk negara bagian Brasil dan wilayah Bahia selatan (lintang -15), Espírito Santo (lintang -18 hingga -22), Rio de Janeiro (lintang -21 hingga -24), São Paulo tengah dan timur laut (lintang -21 hingga -25), Minas Gerais tenggara (lintang -22), Paraná, dan Santa Catarina (lintang -24 hingga -28) (Gambar 2a ).
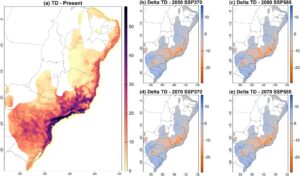
Namun, TD diproyeksikan menurun di masa mendatang, baik dalam skenario optimis maupun pesimis untuk tahun 2050 (Gambar 2b,c ; Gambar S5 ) dan 2070 (Gambar 2d,e ; Gambar S5 ). Wilayah yang diproyeksikan mengalami kehilangan TD tertinggi berada di tenggara AF, termasuk berbagai wilayah yang diproyeksikan memiliki kekayaan tertinggi saat ini. Wilayah yang diproyeksikan mengalami peningkatan TD yang lebih tinggi berada di selatan AF, terutama di negara bagian Rio Grande do Sul dan Santa Catarina di Brasil (lintang -27 hingga -34).
Karena potensi area distribusi spesies sedikit berkurang, ada juga pengurangan PD mamalia kecil (Gambar 3b–e ; Gambar S6 ). Model kami memproyeksikan bahwa wilayah dengan TD tinggi saat ini dan di masa depan kongruen dengan wilayah PD tinggi, seperti yang diharapkan. Kehilangan PD yang tinggi diproyeksikan untuk negara bagian Brasil di Bahia selatan (lintang -13), Espírito Santo (lintang -17 hingga -21), Rio de Janeiro (lintang -21 hingga -23), São Paulo tenggara (lintang -20 hingga -25), Minas Gerais (lintang -17 hingga -22), dan Paraná (lintang -22 hingga -26) (Gambar 3 ).
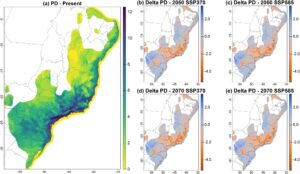
Di sisi lain, beberapa wilayah AF diproyeksikan akan meningkatkan PD di masa mendatang, baik dalam skenario optimistis maupun pesimistis dan untuk tahun 2050 dan 2070. Model memproyeksikan perolehan PD di wilayah utara, tengah, dan selatan AF, termasuk Bahia (lintang -10 hingga -15), utara Minas Gerais (lintang -15 hingga -18), Paraná, Santa Catarina, dan Rio Grande do Sul (lintang -23 hingga -34; Gambar 3 ). Kami juga memproyeksikan area penting untuk konservasi mamalia kecil (area dengan PD SES tinggi dan PD teramati tinggi, Gambar S7 ) di bagian tengah, tenggara, dan timur laut AF untuk saat ini dan masa mendatang. Beberapa kawasan kritis untuk konservasi PD mamalia kecil ini secara spasial bertepatan dengan kawasan yang diproyeksikan mengalami peningkatan TD di masa mendatang, seperti negara bagian Bahia Selatan di Brasil (Garis Lintang -15 hingga -18), São Paulo (Garis Lintang -20 hingga -25), Paraná, dan Mato Grosso do Sul (Garis Lintang -20 hingga -26, Gambar S7 dan Gambar 2 ).
Penurunan PD mamalia kecil khususnya memprihatinkan ketika hanya berfokus pada kawasan lindung dan sisa hutan AF (Gambar 4 ). Model kami menunjukkan bahwa sebagian besar kawasan lindung di sepanjang wilayah tengah dan utara AF diproyeksikan mengalami kehilangan PD mamalia kecil pada tahun 2070 dalam skenario pesimistis, dengan beberapa pengecualian (Gambar 4a ). Di wilayah selatan AF, tempat peningkatan PD terbesar diharapkan, kami mengamati beberapa kawasan lindung kecil dan berjauhan secara spasial (Gambar 4a ). Di sisi lain, kawasan dengan jumlah vegetasi asli yang tinggi dan jumlah kawasan lindung yang lebih tinggi, seperti Serra do Mar di tengah-selatan AF (lintang -22 hingga -26), menunjukkan kehilangan PD yang nyata dalam skenario pesimistis 2070.
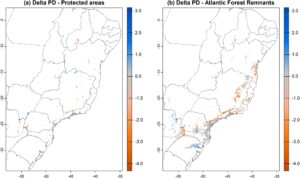
4 Diskusi
Model kami menunjukkan bahwa perubahan dalam suhu dan rezim presipitasi yang diukur dengan variabel bioklimatik cenderung mengurangi taksonomi dan PD mamalia kecil di beberapa wilayah AF secara moderat. Meskipun beberapa wilayah di selatan AF diproyeksikan akan mendapatkan PD di masa mendatang, sebagian besar wilayah ini berada di wilayah dengan sedikit sisa hutan dan kawasan lindung yang terisolasi secara spasial. Sepengetahuan kami, masih belum ada studi yang mengevaluasi pentingnya sisa hutan dan kawasan lindung bagi mamalia kecil di bawah perubahan iklim. Berdasarkan hasil yang ditemukan di sini, kami mengidentifikasi wilayah yang diproyeksikan mengalami kerugian minimum hingga sedang dalam taksonomi dan PD di masa mendatang. Yang lebih penting lagi, model kami memproyeksikan wilayah dengan stabilitas tinggi (SES PD, PD, dan TD tinggi) di mana keanekaragaman dapat dijaga dan harus diprioritaskan untuk konservasi saat ini.
Pola proyeksi TD yang diamati saat ini sesuai dengan penelitian lain yang telah dilakukan dengan mamalia di Amerika Selatan, yang telah menunjukkan TD tinggi mamalia kecil di sepanjang selatan negara bagian Bahia di Brasil, Espírito Santo, Rio de Janeiro, tengah dan timur laut São Paulo, timur laut Minas Gerais, dan beberapa wilayah di utara dan tengah Paraná dan Santa Catarina (Gambar 1 ) (misalnya, Maestri dan Patterson 2016 ; da Silva et al. 2012 ; Costa et al. 2000 ). Selain itu, wilayah-wilayah yang diproyeksikan menyajikan TD tinggi ini secara spasial bertepatan dengan daerah endemisme Caparaó dan Tenggara untuk mamalia kecil yang tidak aktif di AF (Dalapicolla et al. 2021 ). TD tinggi yang diproyeksikan di lokasi-lokasi ini juga didukung oleh penelitian lain yang melibatkan mamalia berukuran sedang dan besar, anuran, dan burung (de Oliveira et al. 2023 ; Vale et al. 2018 ; Vasconcelos et al. 2018 ; Tonetti et al. 2022 ). Selain itu, wilayah-wilayah ini adalah rumah bagi spesies unik yang tidak dapat ditemukan di tempat lain, yang menyoroti pentingnya mereka sebagai hotspot endemisme. Beberapa wilayah ini saat ini menampung spesies yang terancam tingkat tinggi, terutama antara negara bagian Paraná dan São Paulo (IUCN 2024 ), yang diproyeksikan akan sangat rentan terhadap kepunahan lokal. Lebih jauh lagi, banyak spesies di area-area ini kurang diteliti, yang berpotensi menyembunyikan tingkat ancaman dan kerentanan yang lebih tinggi (IUCN 2024 ; Lacher et al. 2020 ).
Model kami menunjukkan bahwa banyak wilayah yang terletak di dalam pusat endemisme diproyeksikan akan kehilangan TD di masa mendatang. Di antara pusat-pusat ini, pusat endemisme Bahia, Caparaó dan Espinhaço (lihat Gambar S4 ) menonjol karena kehilangan TD yang besar pada tahun 2050 dan 2070. Perubahan signifikan dalam TD juga diproyeksikan untuk Tenggara pusat (lihat Gambar S4 ), yang terletak di wilayah Pegunungan Pesisir di Brasil Tenggara. Meskipun wilayah ini, yang dibatasi terutama oleh Serra do Mar, menyajikan persentase besar sisa-sisa hutan dan banyak kawasan lindung, perubahan yang diproyeksikan dalam suhu dan rezim curah hujan kemungkinan akan menyebabkan pengurangan yang kuat di daerah iklim yang cocok untuk mamalia kecil di wilayah ini. Proyeksi pengurangan potensi wilayah sebaran spesies sebagai respons terhadap perubahan iklim juga terjadi pada kelompok lain seperti mamalia (Levinsky dkk. 2007 ; Maiorano dkk. 2011 ; Schloss dkk. 2012 ; Hidasi-Neto dkk. 2019 ), anuran (Alves-Ferreira, Giné, dkk. 2022 ; Alves-Ferreira, Talora, dkk. 2022 ; Anunciação dkk. 2023 ), reptil (Biber dkk. 2023 ), dan burung (Mota dkk. 2022 ; Tonetti dkk. 2022 ).
Penyusutan dalam potensi distribusi spesies dapat menyebabkan pengurangan atau realokasi PD mamalia. Faktanya, perubahan iklim diproyeksikan akan mempengaruhi seluruh klade pohon filogenetik (González-Orozco et al. 2016 ). Model kami menunjukkan bahwa PD mamalia kecil diproyeksikan mengalami penurunan drastis, terutama di tenggara AF. Wilayah ini diproyeksikan mengalami peningkatan besar dalam kejadian ekstrem seperti kekeringan, hujan lebat, banjir, dan tanah longsor (Magrin et al. 2014 ; Castellanos et al. 2022 ). Karena suhu menentukan distribusi mamalia kecil (Maestri dan Patterson 2016 ), ketidakstabilan iklim ini dapat menyebabkan penurunan taksonomi dan PD kelompok vertebrata ini. Studi telah menunjukkan bahwa mamalia dengan jumlah kerabat dekat filogenetik yang rendah lebih mungkin terkena dampak negatif oleh perubahan iklim dibandingkan dengan spesies dengan jumlah spesies yang tinggi dalam genus dan familinya (Russell dan Schupp 1998 ; Purvis et al. 2000 ). Hal ini karena spesies yang berkerabat dekat cenderung memiliki kebutuhan lingkungan dan kerentanan yang sama terhadap ancaman lokal (Jono dan Pavoine 2012 ). Namun, memiliki banyak spesies yang mirip tidak akan selalu mengonfirmasi lebih banyak perlindungan dan/atau lebih sedikit kerentanan, karena beberapa spesies menunjukkan kecenderungan genetik terhadap kepunahan lokal dan sifat-sifat khusus yang meningkatkan risiko kepunahan mereka (Jono dan Pavoine 2012 ).
Skenarionya menjadi lebih buruk ketika kita mempertimbangkan bahwa sebagian besar kawasan lindung dan sisa-sisa AF tidak dapat melindungi PD yang diproyeksikan dari mamalia kecil di bawah skenario perubahan iklim. Sebagian besar sisa-sisa dan kawasan lindung diperkirakan akan mengalami penurunan signifikan dalam PD di masa mendatang. Misalnya, kawasan endemisme di bagian tenggara AF (lihat Gambar S3 ) memusatkan kehilangan PD tertinggi dan layak mendapat perhatian lebih lanjut. Di sisi lain, bagian timur negara bagian Bahia memegang sejumlah besar tutupan hutan dan diperkirakan akan menahan PD di masa mendatang, sehingga mewakili wilayah yang menjanjikan untuk memperluas jaringan kawasan lindung. Keuntungan dalam PD ini dikuatkan oleh pola pergantian yang telah terdeteksi di wilayah yang sama (Maestri dan Patterson 2016 ). Bagian selatan AF juga diproyeksikan mengalami peningkatan PD.
Model kami menunjukkan bahwa daerah dengan garis keturunan generik, suku, dan subfamili yang lebih sedikit, seperti bagian selatan dan timur laut, mungkin mengalami kolonisasi di masa depan oleh spesies yang secara filogenetik berbeda, yang mengarah pada peningkatan PD. Daerah-daerah ini dapat digunakan sebagai tempat perlindungan bagi mamalia kecil dan diharapkan untuk memusatkan nilai-nilai keragaman filogenetik yang tinggi di masa depan. Tempat-tempat ini sangat penting untuk konservasi karena mereka menyoroti daerah yang memiliki warisan evolusi yang tinggi (González-Orozco et al. 2016 ). Selain itu, stabilitas daerah-daerah ini selaras dengan pola historis yang diamati di AF, di mana daerah yang stabil telah bertindak sebagai tempat perlindungan, melestarikan spesies endemik dan garis keturunan peninggalan (Keppel et al. 2012 ; Lourenço-de-Moraes et al. 2019 ). Misalnya, AF pusat berfungsi sebagai tempat perlindungan bagi spesies Neotropis selama akhir Pleistosen (Carnaval et al. 2009 ), dan wilayah dengan elevasi tinggi diharapkan menyediakan tempat perlindungan serupa dalam skenario iklim mendatang (Lemes dan Loyola 2013 ; Struebig et al. 2015 ). Lebih jauh lagi, wilayah yang tidak dilindungi dengan keanekaragaman arus yang lebih rendah dapat berfungsi sebagai koridor untuk migrasi spesies ke habitat baru yang sesuai (Littlefield et al. 2017 , 2019 ). Untuk memastikan kelangsungan hidup mamalia kecil dan mencapai tujuan konservasi jangka panjang, penting untuk tidak hanya menciptakan wilayah perlindungan baru tetapi juga menerapkan langkah-langkah efektif untuk mengurangi penggundulan hutan, kebakaran, dan invasi spesies di wilayah selatan dan timur laut Hutan Atlantik (Watson et al. 2013 ; Borges dan Loyola 2020 ). Tindakan segera diperlukan untuk memulihkan ekosistem, meningkatkan pengelolaan lahan, dan mengamankan keanekaragaman hayati sambil menangani lahan pribadi yang tidak mematuhi undang-undang Brasil (Girardin et al. 2021 ; Rezende et al. 2018 ; Vancine et al. 2024 ).
Kita harus mempertimbangkan bahwa distribusi potensial yang diproyeksikan dalam studi ini mengandung beberapa sumber ketidakpastian. Misalnya, model SDM berasumsi bahwa spesies berada dalam keseimbangan dengan lingkungannya, relung dipertahankan dari waktu ke waktu, dan spesies tidak akan mampu beradaptasi dengan perubahan iklim (Austin 2007). Asumsi-asumsi ini mengabaikan kapasitas spesies untuk bertahan hidup melalui plastisitas atau mengalihkan distribusinya ke wilayah lain ketika dihadapkan dengan kondisi lingkungan yang baru (Boutin dan Lane 2014 ; Santos et al. 2017 ). Meskipun demikian, karena persyaratan ekologis dan plastisitas rendah yang diharapkan dari mamalia kecil, terutama yang menghuni lanskap yang diubah oleh antropogenik, pergeseran distribusi dan adaptasi cepat merupakan strategi terbatas bagi spesies ini untuk secara efektif menanggapi perubahan iklim yang cepat (Boutin dan Lane 2014 ). Lebih jauh lagi, S-SDM sangat rentan terhadap kritik dan mungkin tidak mewakili komposisi spesies, sering kali melebih-lebihkan komposisi spesies (Calabrese et al. 2014 ; Zwiener et al. 2023). Meskipun demikian, studi kami sejalan dengan studi lain yang telah dipublikasikan untuk beberapa kelompok taksonomi yang menyimpulkan dampak perubahan iklim pada AF (misalnya, Esser et al. 2019 ; Santos et al. 2020 ; Tonetti et al. 2022 ; Anunciação et al. 2023 ; Ribeiro-Souza et al. 2024 ). Akan tetapi, studi mendatang mungkin mempertimbangkan keterbatasan ini dan menerapkan metode yang lebih kuat untuk memperkirakan komposisi spesies (misalnya, Guisan dan Rahbek 2011 ; Calabrese et al. 2014 ).
Sebagai kesimpulan, model kami menunjukkan bahwa perubahan iklim kemungkinan besar akan berdampak signifikan pada komunitas mamalia kecil di AF. Kenaikan emisi gas rumah kaca yang diusulkan, meskipun sedang, diperkirakan akan mengurangi kesesuaian iklim untuk mamalia kecil di seluruh AF, sehingga menyusutkan wilayah dengan kekayaan spesies yang tinggi. Peningkatan suhu rata-rata yang substansial akan mengintensifkan penurunan di wilayah yang mendukung iklim bagi mamalia kecil (studi ini), mamalia sedang dan besar bahkan lebih jauh (de Oliveira et al. 2023 ). Secara khusus, model kami memproyeksikan bahwa wilayah tenggara AF kemungkinan akan mengalami penurunan PD yang paling nyata, sementara beberapa wilayah di selatan dapat berfungsi sebagai tempat perlindungan iklim bagi spesies ini. Namun, kawasan lindung dan sisa hutan saat ini diproyeksikan tidak akan melindungi keanekaragaman mamalia kecil di bawah perubahan iklim di masa mendatang. Studi kami menyoroti kebutuhan mendesak untuk tindakan guna mengurangi dampak perubahan iklim pada mamalia kecil dan melindungi keanekaragaman filogenetiknya. Kami merekomendasikan agar upaya konservasi mempertimbangkan wilayah selatan AF sebagai wilayah target, karena wilayah ini diproyeksikan akan memperoleh TD dan PD di masa mendatang. Wilayah-wilayah ini dapat menjadi penting untuk menjaga sejarah evolusi mamalia kecil. Mengatasi masalah ini dapat menjadi langkah awal untuk melindungi keanekaragaman hayati dan menjaga sejarah evolusi komunitas mamalia kecil terhadap dampak perubahan iklim yang tak terelakkan (Bellard et al. 2012 ).