
ABSTRAK
Gangguan antropogenik telah lama diakui sebagai salah satu ancaman utama bagi keanekaragaman hayati di seluruh dunia; namun, masih sedikit yang dipahami tentang bagaimana infrastruktur yang dibangun manusia memengaruhi aliran gen dan struktur filogeografi tumbuhan. Informasi tersebut berguna untuk konservasi dan pemulihan ekosistem yang terganggu manusia. Di sini kami mempelajari dampak reservoir besar dan dua jalan tol pada Primulina heterotricha (Gesneriaceae), herba berumur pendek yang endemik di Pulau Hainan (Tiongkok), salah satu area utama hotspot keanekaragaman hayati Indo-Burma yang penting secara global. Dengan menerapkan filogeografi komparatif menggunakan satu DNA ribosom nuklir dan dua sekuens DNA kloroplas, kami memperkirakan tingkat keanekaragaman genetik dan diferensiasi pada 176 dan 117 individu yang dikumpulkan, masing-masing, sebelum (tahun 2016) dan setelah (tahun 2022) pembangunan dua jalan tol di Pulau Hainan, dari delapan populasi P. heterotricha yang sama . Kami menemukan bahwa spesies ini secara signifikan meningkatkan diferensiasi genetik nuklir selama periode 2016-2022, yang bertepatan dengan pembukaan dua jalan bebas hambatan tersebut. Yang juga perlu diperhatikan, pembagian ribotipe di antara tiga kelompok populasi yang dipisahkan oleh jaringan jalan bebas hambatan tersebut sangat berkurang selama periode yang sama. Selain itu, perubahan dalam signifikansi hambatan genetik sebelum dan sesudah pembangunan jalan menunjukkan bahwa isolasi geografis yang disebabkan oleh pembangunan manusia adalah kunci untuk memahami pola filogeografis P. heterotricha saat ini . Kami memberikan bukti langsung bahwa infrastruktur antropogenik yang besar mampu meningkatkan diferensiasi genetik dan, dengan demikian, memodifikasi pola filogeografis P. heterotricha , hanya dalam periode enam tahun (atau dua generasi tanaman studi). Kami menyarankan untuk membangun koridor ekologis guna meningkatkan pertukaran gen antara kedua sisi hambatan buatan ini.
1 Pendahuluan
Aktivitas manusia, yang dianggap sebagai ancaman utama bagi keanekaragaman hayati di seluruh dunia, mampu mengganggu integritas habitat dan, dengan demikian, memodifikasi pola filogeografi spesies yang terdistribusi secara alami (Young et al. 1996 ; Aguilar et al. 2006 ; Martínez-Ramos et al. 2016 ). Misalnya, infrastruktur antropogenik seperti jalan bebas hambatan, gedung, dan lahan pertanian yang luas dapat mengganggu penyerbukan tanaman dan penyebaran benih dengan menghambat pergerakan serangga dan hewan lainnya (Honnay dan Jacquemyn 2007 ). Fragmentasi populasi alami dan isolasi geografis individu juga dapat menyebabkan peningkatan tingkat perkawinan sedarah dan pergeseran genetik (Lowe et al. 2015 ), yang menempatkan populasi pada risiko kepunahan (Tambarussi et al. 2017 ; Moraes et al. 2018 ).
Pulau Hainan, yang terletak di Cina Selatan (Gambar 1 ), memiliki luas 35.000 km 2 dan merupakan bagian khas dari hotspot keanekaragaman hayati Indo-Burma yang penting secara global (Myers et al. 2000 ), karena menampung sekitar 4800 tanaman vaskular secara total dengan hampir 500 endemik (Francisco-Ortega et al. 2010a , 2010b ; Yang 2013 ). Sebagian besar keanekaragaman tumbuhan yang kaya ini (misalnya, 80% spesies endemik) terkonsentrasi di sistem pegunungan selatan-tengah pulau (Francisco-Ortega et al. 2010a , 2010b ). Khususnya, daerah ini adalah rumah bagi primata paling terancam punah di dunia Nomascus hainanus (Long et al. 2021 ), yang menyoroti nilai konservasinya.
Namun, saat ini, Pulau Hainan tengah mengalami perkembangan ekonomi yang pesat, yang sebagian didasarkan pada pembangunan infrastruktur besar; misalnya, pemerintah setempat tengah berupaya membangun pelabuhan perdagangan bebas terbesar di dunia ( www.xinhuanet.com/english/2018-04/13/c_137109243.htm ). Bentang alam dan vegetasi telah sangat terpengaruh selama beberapa dekade terakhir akibat percepatan pertumbuhan populasi manusia dan urbanisasi. Misalnya, Waduk Daguangba di Sungai Changhua, sungai terbesar kedua di pulau itu, selesai dibangun pada tahun 1994 dan membentuk badan air lamban dengan lebar hampir 6 km. Waduk ini, yang terletak di antara Gunung E’xian dan Gunung Jianfeng, dapat mengganggu konektivitas vegetasi alami kedua gunung ini. Selain itu, dua jalan tol baru-baru ini dibangun: Jalan G9811 (selanjutnya disebut Jalan Tol 1), yang selesai dibangun pada tahun 2018 dan memiliki lebar 26 m, dan Jalan S10 (selanjutnya disebut Jalan Tol 2), yang dibangun pada tahun 2019 dan memiliki lebar 20 m ( www.xinhuanet.com/english/2021-04/15/c_139882827.htm ). Konstruksi antropogenik ini terletak di bagian tengah pegunungan selatan-tengah di pulau tersebut dan dapat memengaruhi integritas habitat dan dinamika populasi tanaman langka dan terancam punah yang ada di bagian pulau ini, yang selanjutnya dapat mengakibatkan perubahan dalam struktur filogeografinya.
Famili Gesneriaceae di Pulau Hainan dicirikan oleh tingkat keanekaragaman spesies dan endemisme yang tinggi (Wei 2010 ; Ling et al. 2017a , 2017b ). Sebanyak empat belas genus dan 25 spesies famili ini hidup di pulau tersebut, termasuk satu genus endemik dan 10 spesies endemik (Li dan Wang 2005 ; Yang 2013 ; Ling et al. 2017a ; Ling, Guan, et al. 2020 ). Primulina heterotricha (Merr.) Y. Dong & Yin Z. Wang (Gambar 1 ), salah satu spesies endemik ini, tersebar luas di pegunungan selatan-tengah pulau (Ling et al. 2017a , 2017b ), yang mungkin disebabkan, antara lain, oleh kemampuannya tumbuh di tanah asam dan basa (Ai et al. 2015 ) Tumbuhan, herba berumur pendek (dengan waktu generasi 1–3 tahun), memiliki bunga tubular zygomorfik yang diserbuki oleh beberapa serangga, terutama Glossamegilla malaccensis dan G. yunnanensis dari Anthophoridae (Ling et al. 2017a , 2017b ). Buah (kapsul) tegak dengan biji kecil berwarna cokelat dan berbentuk fusiform, menunjukkan kemampuan penyebaran yang buruk yang kemungkinan terkait dengan aliran air (MXR, pengamatan pribadi). Primulina heterotricha merupakan tanaman diploid dengan 2 n = 36 (Kang et al. 2014 ). Dengan mempertimbangkan semua faktor yang disebutkan di atas (yaitu, distribusinya yang cukup berkesinambungan di area penelitian, waktu generasinya yang singkat, dan kemampuan penyebarannya yang terbatas), spesies ini merupakan sistem model yang baik untuk mengungkap kemungkinan efek fragmentasi jangka pendek dari Waduk Daguangba dan jalan tol yang baru dibangun terhadap diferensiasi genetik dan pola filogeografinya.
Dalam studi ini, kami membahas pertanyaan-pertanyaan berikut: (1) Apakah waduk dan jalan tol yang baru dibangun (yaitu, penghalang buatan) sudah memengaruhi susunan genetik P. heterotricha endemik Hainan ? dan (2) jika ya, bagaimana penghalang buatan ini memengaruhi diferensiasi populasi dan pola filogeografi spesies yang diteliti? Untuk mendapatkan wawasan tentang pertanyaan-pertanyaan ini, kami melakukan genetika populasi dan studi filogeografi berdasarkan sampel P. heterotricha yang dikumpulkan pada tahun 2016 dan 2022 (yaitu, sebelum dan sesudah pembangunan jalan tol), menggunakan urutan DNA nuklir dan kloroplas yang terkonservasi dan berevolusi lambat.
2 Bahan dan Metode
2.1 Pengumpulan Sampel dan Prosedur Laboratorium
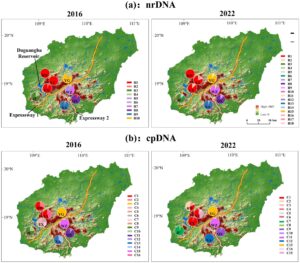
Delapan populasi yang secara kasar mencakup seluruh rentang distribusi P. heterotricha diambil sampelnya dan meliputi BW (Gunung Bawang), YJ (Gunung Yajia), EX (Gunung Erxian), YG (Gunung Yingge), QX (Gunung Qixian), WZ (Gunung Wuzhi), XA (Hutan Batu Xian’an), dan JF (Gunung Jianfeng) (Tabel 1 , Gambar 2 ). Sebanyak 176 sampel dikumpulkan pada tahun 2016 dan 117 sampel pada tahun 2022 (Tabel 1 ). Meskipun kami tidak mengambil sampel sebelum tahun 1994 ketika Waduk Daguangba di Sungai Changhua selesai dibangun, pengambilan sampel dari delapan populasi sebelum (2016) dan setelah (2022) pembangunan dua jalan tol (jalan tol 1 dan 2 masing-masing selesai dibangun pada tahun 2018 dan 2019) akan memungkinkan kami untuk mendeteksi kemungkinan efek genetik dan filogeografi dari fragmentasi habitat. Jarak antara populasi berkisar dari 6 hingga 72 km, dan ketinggian populasi yang dikumpulkan bervariasi dari 600 (YG) hingga 1163 m (YJ) di atas permukaan laut (Tabel 1 ). Mempertimbangkan hambatan fisik yang ditimbulkan oleh dua jalan bebas hambatan dan bendungan, populasi dapat dibagi sementara menjadi empat kelompok: kelompok populasi barat laut (NW) (yaitu, BW, YJ, EX, dan YG), kelompok tenggara (SE) (QX dan WZ), kelompok selatan (S) (XA), dan kelompok barat daya (SW) (JF). Pada setiap populasi, sampel daun segar dikumpulkan dari individu yang berjarak setidaknya 10 m. Setiap daun yang diambil sampelnya dikeringkan dengan cepat dalam kantong plastik terpisah yang berisi 20 hingga 30 g gel silika dan disimpan pada suhu −80°C kemudian.
| Lokasi pengambilan sampel | Kode populasi | Nomor voucher | Ketinggian (m) | Ukuran sampel | |
|---|---|---|---|---|---|
| Tahun 2016 | Tahun 2022 | ||||
| Gunung Bawang | BW | Ling20160803PH003 | tahun 1011 | 16 | 16 |
| Gunung Yajia | YJ | Ling20160807PH011 | tahun 1163 | 16 | 15 |
| Gunung Exian | MANTAN | Ling20160807PH019 | 920 | 31 | 16 |
| Gunung Yingge | YG | Ling20160809PH005 | 600 | 9 | 10 |
| Gunung Qixian | QX | Ling20160809PH013 | 1000 | 30 | 15 |
| Gunung Wuzhi | WZ | Ling20161003PH102 | tahun 1058 | 25 | 15 |
| Gunung Xianan | Bahasa Inggris: XA | Ling20161015PH017 | 783 | 16 | 15 |
| Gunung Jianfeng | JF | Ling20161017PH009 | 918 | 33 | 15 |
| Jumlah | 176 | 117 | |||
Total DNA genomik untuk semua sampel diekstraksi menggunakan prosedur standar CTAB (Doyle dan Doyle 1987 ) dari 30 mg jaringan daun kering dan dijadikan cetakan untuk reaksi berantai polimerase (PCR). Kualitas dan kuantitas DNA ditentukan pada gel agarosa 0,8% yang diwarnai dengan 2,5 μL Goldview (Aidlab Biotechnologies Co. Ltd., Beijing, Tiongkok), dengan penanda DNA AL2000 (Aidlab Biotechnologies).
Satu urutan DNA ribosom nuklir (nrDNA), daerah spacer inter-transkripsi ribosom (ITS) yang terdiri dari spacer 1, gen ribosom 5.8S, dan spacer 2 (White et al. 1990 ), dan dua daerah spacer intron DNA kloroplas (cpDNA) trn L- trn F (Taberlet et al. 1991 ) dan ycf 1b (Dong et al. 2015 ) digunakan dalam penelitian ini (Tabel 2 ). Kami menggunakan urutan nuklir dan kloroplas karena pewarisan mereka yang berbeda (biparental untuk nrDNA dan maternal untuk cpDNA di sebagian besar angiospermae) memfasilitasi pemahaman efek aliran gen yang dimediasi biji dan serbuk sari pada struktur genetik, sementara penggunaan gabungan mereka umumnya menghasilkan inferensi filogenetik yang lebih kuat. Kami telah menambahkan ycf 1b ke kombinasi berulang ITS/ trn L- trn F dalam genetika tanaman karena merupakan salah satu wilayah yang paling bervariasi pada tanaman (Dong et al. 2015 ).
| Fragmen DNA | DIA | trn L-F dan ycf 1b | ||
|---|---|---|---|---|
| Tahun 2016 | Tahun 2022 | Tahun 2016 | Tahun 2022 | |
| S | 13 | 26 | 48 | 42 |
| Tidak | 10 | 18 | 16 | 15 |
| H | 0.809 | 0.876 | 0.907 | 0,874 tahun |
| π | 0,00435 | 0,00464 tahun | 0,00940 | 0,00868 |
| Tidak ada | 2.98812 | 2.91217 | 12.72468 | 13.63660 |
| Ukuran fragmen | 701 | 640 | tahun 1405 | tahun 1592 |
| D dari Tajimas | 0.80474 | -1.18345 | 1.38922 | 2.25892 |
| F – nya Fu | 1.811 | -3.937 | 12.439 | 11.3 |
Reaksi PCR disiapkan dalam volume 25 μL, yang terdiri dari 20 μL ddH 2 O, 2,5 μL buffer 10 ×, 0,5 μL dNTP 10 mM, 0,5 μL masing-masing primer 5 μM, 0,5 μL cetakan DNA (sekitar 0,1 ng) dan 0,5 μL polimerase Taq 5 U/μL (Aidlab Biotechnologies). Reaksi PCR dilakukan pada Thermal Cycler 2720 (Applied Biosystems oleh Life Technologies, Singapura). Program PCR untuk ITS1/2 dan trn L- trn F dirancang dengan denaturasi awal 5 menit pada suhu 94°C, diikuti oleh 35 siklus 1 menit pada suhu 94°C, 1 menit pada suhu 55°C, 1 menit pada suhu 72°C, dan ekstensi akhir 10 menit pada suhu 72°C. Amplifikasi ycf 1b menggunakan protokol berikut: 4 menit pada suhu 94°C, 35 siklus 30 detik pada suhu 94°C, 40 detik pada suhu 58°C, dan 1 menit pada suhu 72°C, diakhiri dengan 10 menit pada suhu 72°C. Semua produk PCR diverifikasi berdasarkan ukuran dengan elektroforesis gel. Amplikon diurutkan oleh ABI 3730 DNA Analyzer dalam arah maju dan mundur berdasarkan BigDye Terminator Cycle Sequencing Ready Kit (Applied Biosystems, Foster City, CA, AS) di BGI (Beijing Genomics Institute, Cina).
2.2 Keanekaragaman Genetik
Kromatogram dari kedua arah sekuens ITS1/2 dan cpDNA diperiksa secara visual dan diedit secara manual dengan perangkat lunak BioEdit (Hall 1999 ) untuk konfirmasi basa dan pengeditan sekuens yang berdekatan; setiap basa/karakter diberi bobot yang sama sebelum analisis, dan setiap indel/gap direpresentasikan sebagai mutasi tunggal. Tiga sekuens disejajarkan secara manual dan dipangkas secara integral jika perlu menggunakan MEGA v.6.5 (Kumar et al. 2008 ) secara terpisah. Dua sekuens cpDNA non-coding dirakit sebagai lokus tunggal untuk analisis selanjutnya oleh SequenceMatrix v.1.7.8 (Vaidya et al. 2011 ), dan uji homogenitas partisi sekuens cpDNA vs. sekuens nrITS dilakukan dengan PAUP* v.4.0a164 (Swofford 2002 ). Karena ketidakhomogenan kedua matriks terdeteksi, nrITS dan cpDNA dianalisis secara independen.
DNASP v.6.12.01 (Rozas et al. 2017 ) digunakan untuk menghitung jumlah ribotipe/klorotipe yang teridentifikasi ( Nn ), keanekaragaman haplotipe ( h ) dalam populasi, situs polimorfik ( S ), keanekaragaman nukleotida ( π ), dan jumlah rata-rata perbedaan nukleotida ( Nd ) secara terpisah untuk matriks nrITS dan cpDNA. Uji Mann–Whitney U (yang dilakukan dengan “ Kalkulator Uji Mann-Whitney U ” yang tersedia di https://www.socscistatistics.com/tests/mannwhitney/ ) digunakan untuk menilai signifikansi perbedaan dalam Nn , h , S , dan π antara tahun 2016 dan 2022. Peta distribusi geografis ribotipe/klorotipe pada tingkat populasi, juga secara terpisah untuk sekuens nrITS dan cpDNA, dibuat dan divisualisasikan dengan ArcGIS v.10.8 (ESRI, Redlands, CA, AS).
2.3 Hubungan Filogenetik
Kami menyimpulkan model optimal substitusi nukleotida menggunakan MrModeltest v.2.3 (Nylander 2004 ), berdasarkan AIC (Akaike Information Criteria) (Akaike 1981 ). Model yang paling sesuai (GTR + I + G) digunakan dalam menyimpulkan hubungan filogenetik ribotipe yang diidentifikasi, yang dilakukan melalui pohon kemungkinan maksimum (ML) dan inferensi Bayesian (BI) (Gao et al. 2015 ; Genbank dengan nomor akses DQ872827 ).
Analisis ML ribotipe dilakukan menggunakan MEGA v.6.5 dengan model substitusi optimal dan 1000 replikasi bootstrap untuk menilai dukungan kelompok yang dihasilkan. Analisis BI ribotipe dilakukan menggunakan MrBayes v.3.2.6 (nst = 2, rates = equal) (Ronquist et al. 2012 ), menggunakan model substitusi nukleotida optimal yang disimpulkan oleh AIC dalam PAUP v.4.0a164. Analisis dimulai dengan pohon acak menggunakan rantai Markov Chain Monte Carlo (MCMC) dengan 10 juta generasi, mengambil sampel setiap 10.000 generasi di empat operasi Bayesian independen. 2500 pohon pertama (25% dari total pohon) dibuang sebagai burn-in, dan pohon yang tersisa diringkas dalam pohon konsensus aturan mayoritas 50% dengan probabilitas posterior (PP). Konvergensi rantai dinilai dengan memeriksa ukuran sampel efektif yang lebih besar dari 200 untuk setiap parameter di Tracer v.1.6 (Rambaut dan Drummond 2007 ), dan panjang dan PP setiap cabang divisualisasikan oleh FIGTREE v.1.4.2 (Rambaut 2009 ).
2.4 Struktur Populasi, Analisis Distribusi Ketidaksesuaian, dan Deteksi Netralitas
Struktur populasi dari urutan nrDNA dan cpDNA disimpulkan menggunakan prosedur pengelompokan Bayesian yang diterapkan dalam STRUCTURE v.2.3.4 (Evanno et al. 2005 ) tanpa informasi struktur sebelumnya; perangkat lunak ini mengidentifikasi jumlah yang paling mungkin ( K ) dari klaster genetik asal individu yang dijadikan sampel dan menetapkan individu ke dalam klaster. Kami menggunakan iterasi MCMC sebagaimana diterapkan dalam STRUCTURE untuk mengeksplorasi ruang parameter dengan mempertimbangkan keanggotaan individu ke klaster K , mulai dari K = 1 (hipotesis nol panmixia) hingga K = 8 (jumlah total populasi yang dijadikan sampel). Tiga kali pengujian independen dilakukan dengan model campuran pada 10 5 iterasi MCMC dan periode burn-in 10 5 . Jumlah kelompok yang paling mungkin ( K , yang menunjukkan jumlah klaster sebenarnya dalam data) dan nilai model (Δ K , menurut tingkat perubahan orde kedua klaster K yang paling sesuai dengan data) dihitung dalam STRUCTURE HARVESTER (Earl dan vonHoldt 2012 ). Representasi grafis dari hasil dilakukan di server CLUMPAK ( http://clumpak.tau.ac.il/index.html ).
Analisis varians molekuler (AMOVA) dilakukan pada urutan nrDNA dan cpDNA secara terpisah untuk menguji diferensiasi genetik dalam populasi, di antara kelompok, dan populasi dalam kelompok menggunakan GenAlEx v.6.503 (Peakall dan Smouse 2012 ). F ST berpasangan populasi diukur menggunakan DNASP v.6.12.01, dan jarak geografis berpasangan populasi dihitung oleh GenAlEx. Uji Mann–Whitney U juga digunakan untuk menentukan apakah ada perbedaan signifikan dalam nilai F ST berpasangan populasi baik untuk nrDNA maupun cpDNA antara tahun 2016 dan 2022. Untuk menguji apakah ada variasi genetik lokal yang disebabkan oleh isolasi berdasarkan jarak di antara populasi, estimasi F ST /(1− F ST ) dan logaritma natural jarak geografis yang sesuai (dalam km) antara semua kombinasi berpasangan dari delapan populasi diregresikan dan dikenakan uji Mantel (Mantel 1967 ) dengan 999 permutasi acak dalam GenAlEx.
Hubungan genealogis antara ribotipe/klorotipe berdasarkan model Medium-Joining disimpulkan menggunakan NETWORK v.4.6.1.0 ( http://www.fluxus-Engineering.com/ ). Untuk melakukannya, pertama-tama kami menyimpan data sekuensing sebagai format .rdf di DNASP v.6.12.01, kemudian kami membukanya melalui perangkat lunak NETWORK, dan akhirnya, kami memilih model medium-joining untuk mengeluarkan hasilnya. Untuk mengidentifikasi dan mengukur potensi hambatan genetik di antara populasi P. heterotricha dari dataset nrITS dan cpDNA, kami menghitung algoritma perbedaan maksimum Monmonier di Barrier v.2.2 (Manni et al. 2004 ). Kekokohan hambatan ini dinilai dengan melakukan bootstrapping jarak genetik.
Untuk mendeteksi kemungkinan perluasan jangkauan terkini, Tajima’s D (Tajima, 1989 ) dan Fu’s F s (Fu, 1997 ) dihitung untuk menguji penyimpangan dari hipotesis nol tentang ukuran populasi konstan dan evolusi netral untuk setiap fragmen DNA. Distribusi ketidakcocokan berpasangan dan uji netralitas untuk semua populasi dilakukan dalam DNASP menggunakan dataset nrITS dan cpDNA secara terpisah.
3 Hasil
3.1 Dataset nrITS dan cpDNA
Untuk sampel tahun 2016, matriks sekuens ITS1/2 terdiri dari total 701 bp, mengandung 13 situs polimorfik ( S ) dan 10 ribotipe ( Nn ) dari 176 sampel (Tabel 2 ). Pada tingkat spesies, keragaman nukleotida ITS1/2 ( π ) adalah 0,00435, jumlah rata-rata perbedaan nukleotida ( Nd ) adalah 2,988, dan keragaman haplotipe ( h ) adalah 0,809 (Tabel 2 ). Distribusi geografis ribotipe menunjukkan bahwa R1, R5, dan R6 terjadi pada lebih dari satu populasi. Ribotipe R1 dimiliki oleh BW, YJ, EX, dan QX, R5 dimiliki oleh QX dan WZ, dan R6 dimiliki oleh YG, WZ, dan XA. Tujuh ribotipe lainnya bersifat privat (Tabel 3 ; Gambar 2a ).
| Populasi | Tahun 2016 | Tahun 2022 | ||||
|---|---|---|---|---|---|---|
| Tipe (jumlah individu) | H | Sebuah | Tipe (jumlah individu) | H | π | |
| DIA | ||||||
| BW | R1 (16) | angka 0 | angka 0 | R1 (16) | angka 0 | angka 0 |
| YJ | R1 (16) | angka 0 | angka 0 | Bagian 1 (12), Bagian 16 (1) , Bagian 17 (1) , Bagian 18 (1) | 0.3714 | 0,00115 |
| MANTAN | R1 (29), R2 (2) | 0.1247 | 0,00018 | R1 (14), R2 (2) | 0.2333 | 0,00464 tahun |
| YG | R6 (1), R10 (8) | 0.2222 | 0,00032 | R11 (1), R15 (9) | 0.2000 | 0,00032 |
| QX | Nomor 1 (2), Nomor 5 (1), Nomor 8 (27) | 0.1908 | 0,00064 tahun | R7 (1) , R8 (13) , R9 (1) | 0.2571 | 0,00063 |
| WZ | R5 (13), R6 (2), R7 (10) | 0,5867 tahun | 0,00285 | R10 (13) , R11 (1), R12 (1) | 0.2571 | 0,00127 |
| Bahasa Inggris: XA | Bagian 6 (1), Bagian 9 (15) | 0.1250 | 0,00036 | R13 (14) , R14 (1) | 0.3619 | 0,00464 tahun |
| JF | R3 (27), R4 (6) | 0,3068 tahun | 0,00044 | R3 (11), R4 (1) , R5 (1) , R6 (2) | 0.4667 | 0,00365 |
| Jumlah | 1.5562 | 0,00479 | 2.1475 | 0,0163 | ||
| trn L-F dan ycf1 b | ||||||
| BW | Bagian 1 (11), Bagian 2 (5) | 0.4583 | 0,00033 | Bahasa Indonesia: C1 (5), C2 (8) , C3 (1), C4 (1) , C5 (1) | 0.6833 | 0,0024 jam |
| YJ | Bagian 1 (9), Bagian 2 (1), Bagian 16 (6) | 0,5750 | 0,00082 | C1 (8), C3 (1), C14 (1) , C15 (5) | 0.6381 | 0,00204 |
| MANTAN | Bahasa Indonesia:Garis 4 (15), Garis 5 (2) , Garis 6 (5), Garis 7 (6), Garis 8 (3) | 0.7118 | 0,00380 | C1 (15), C6 (1) | 0.1250 | 0,00868 |
| YG | C3 (9) | angka 0 | angka 0 | C13 (10) | angka 0 | angka 0 |
| QX | Bagian 11 (27) , Bagian 12 (3) | 0.1862 | 0,00027 | C9 (14) , C10 (1) | 0.1333 | 0,00008 |
| WZ | C15 (25) | angka 0 | angka 0 | Bab 11 (15) | angka 0 | angka 0 |
| Bahasa Inggris: XA | Kelas 13 (10), Kelas 14 (6) | 0,5000 | 0,00074 tahun | C12 (15) | angka 0 | angka 0 |
| JF | C9 (25), C10 (8) | 0,3788 | 0,00028 | C7 (14), C8 (1) | 0.1333 | 0,00008 |
| Jumlah | 2.8101 | 0,00624 pukul 0,00624 | 1.713 | 0,01328 | ||
Untuk sampel 2022, matriks sekuens ITS1/2 terdiri dari total 640 bp (Tabel 2 ). Dua puluh enam S hadir, yang memungkinkan identifikasi 18 ribotipe berbeda dari ukuran 117 sampel (Tabel 2 ). Pada tingkat spesies, π adalah 0,00464, Nd adalah 2,912, dan h adalah 0,876 (Tabel 2 ). Tidak ada perbedaan statistik ( p > 0,05) ditemukan antara 2 tahun mengenai S , Nn , h , dan π. Dua ribotipe, R1 dan R11, dibagikan oleh setidaknya dua populasi (R1 hadir dalam BW, YJ, dan EX, dan R11 terjadi pada YG dan WZ). 16 ribotipe lainnya bersifat privat (Tabel 3 , Gambar 2a ). Membandingkan sampel antara tahun 2016 dan 2022, ada penurunan baik dalam jumlah (tiga pada tahun 2016 menjadi dua pada tahun 2022) dan persentase ribotipe yang sama (30,0% pada tahun 2016 dan 11,1% pada tahun 2022) (Tabel 3 ). Khususnya, pembagian ribotipe di antara tiga kelompok populasi yang dipisahkan oleh jaringan jalan bebas hambatan (NW, S, dan SE) sangat berkurang; sementara pada tahun 2016 R1 dan R6 dibagi oleh NW dan SE, dan R6 dibagi oleh NW, S, dan SE, pada tahun 2022 hanya R11 yang dibagi oleh NW dan SE (Gambar 3a ).
Panjang total gabungan penyelarasan kloroplas pada tahun 2016 adalah 1405 bp (berukuran 730 dan 674 bp untuk trn L- trn F dan ycf 1b, berturut-turut). Penyelarasan tersebut mengandung 48 S , dan 16 klorotipe ( Nn ) hadir di antara 176 sampel (Tabel 2 ). Pada tingkat spesies, π adalah 0,00940, Nd adalah 12,725, dan h adalah 0,907 (Tabel 2 ). Dari 16 klorotipe, hanya dua (C1 dan C2) yang dibagi antara populasi (BW dan YJ). 14 klorotipe lainnya bersifat privat (Tabel 3 , Gambar 2b ). Untuk sampel 2022, penyelarasan gabungan dua daerah cpDNA terdiri dari 1592 bp (berukuran 870 dan 722 bp untuk trn L- trn F dan ycf 1b, berturut-turut) yang berisi 42 S , dan 15 Nn yang hadir di antara 117 sampel (Tabel 2 ). Pada tingkat spesies, π adalah 0,00868, Nd adalah 13,637, dan h adalah 0,874 (Tabel 2 ). Sedangkan untuk cpDNA, tidak ada perbedaan statistik ( p > 0,05) antara 2016 dan 2022 mengenai S , Nn , h , dan π. Dari 15 klorotipe, hanya dua (C1 dan C3) yang dibagikan (C1 dibagikan oleh BW, YJ, dan EX, dan C3 oleh BW dan YJ) (Tabel 3 ). Ke-13 klorotipe lainnya bersifat privat (Tabel 3 , Gambar 2b ). Singkatnya, dan bertentangan dengan ITS, tidak ada perbedaan besar antara kumpulan data cpDNA 2016 dan 2022, dengan jumlah yang sama (dua) dan persentase yang hampir sama (12,5% vs. 13,3%) klorotipe bersama, semuanya terjadi pada kelompok populasi NW.
3.2 Hubungan Filogenetik
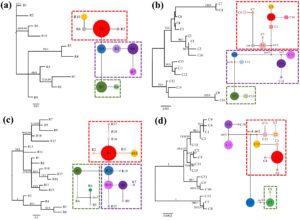
Untuk sampel tahun 2016, jaringan tersebut memisahkan 10 ribotipe ITS nuklir menjadi tiga garis keturunan monofiletik yang berbeda. Jelas, ribotipe R1, R2, R6, dan R10 yang ada dalam kelompok populasi barat laut (NW) membentuk klade; R5 dan R7–R9 yang mendominasi populasi selatan (S) dan tenggara (SE) memiliki hubungan filogenetik yang paling dekat dengan klade NW, sedangkan ribotipe R3 dan R4 yang eksklusif untuk populasi barat daya (SW) membentuk klade independen dari NW (Gambar 3a ). Demikian pula, untuk sampel tahun 2022, jaringan tersebut memisahkan 18 ribotipe ITS nuklir menjadi tiga garis keturunan monofiletik. Ribotipe R1, R2, R11, R15–R17, dan R18, yang mendominasi populasi NW, menyatu menjadi satu kelompok; R7–R10 dan R12–R14 dari kelompok S dan SE menunjukkan hubungan filogenetik yang paling dekat, dan R3–R5 dan R6, yang hanya terdapat pada kelompok SW, membentuk klade yang independen (Gambar 3c ). Selain itu, pohon BI dengan nilai BS/PP yang tinggi berdasarkan ribotipe menunjukkan hasil yang sangat mirip (Gambar S1a,b ).
Mengenai cpDNA, pola hubungan filogenetik sangat dekat dengan nrDNA. Untuk kedua periode pengambilan sampel (2016 dan 2022), jaringan klorotipe menunjukkan tiga garis keturunan monofiletik (Gambar 3b,d ). Klorotipe yang paling melimpah dalam kelompok populasi NW (C1–C8 dan C16 pada tahun 2016; C1–C6, C13, C14, dan C15 pada tahun 2022) membentuk klade, yang paling dekat dengan klade yang dibentuk oleh klorotipe dari kelompok populasi S dan SE (C12–C15 pada tahun 2016, C9–C12 pada tahun 2022); klade yang dibentuk oleh klorotipe eksklusif untuk kelompok SW (populasi JF; C9 dan C10 pada tahun 2016, C7 dan C8 pada tahun 2022; Gambar 3b,d ) adalah yang paling jauh dari yang NW. Sekali lagi, pohon BI dengan nilai BS/PP tinggi berdasarkan klorotipe menunjukkan hasil yang sangat mirip (Gambar S1c,d ).
3.3 Struktur Genetik Populasi dan Pola Filogeografi
Diferensiasi genetik berpasangan ( F ST ) rendah untuk dataset nrITS: untuk sampel tahun 2016, nilai F ST berkisar dari 0 (pasangan BW-EX dan YJ-EX) hingga 0,010 (pasangan JF-YG dan JF-WZ) dengan rata-rata 0,0045; estimasi untuk sampel tahun 2022 berkisar dari 0 (pasangan BW-EX) hingga 0,013 (JF-YG), dengan rata-rata 0,0054 (Tabel S1 ). Mengenai dataset cpDNA, nilai F ST untuk sampel 2016 berkisar dari 0,001 (pasangan BW-YJ) hingga 0,020 (pasangan JF-EX dan JF-YG), dengan rata-rata 0,0098, sementara nilai untuk sampel 2022 berkisar dari 0,001 (pasangan JF-XA) hingga 0,018 (pasangan EX-XA, EX-JF, OX-XA, OX-JF, WZ-XA, dan WZ-JF), rata-rata 0,0097 (Tabel S2 ). Populasi yang dipisahkan dari sisanya oleh bendungan (JF) menunjukkan nilai F ST berpasangan terbesar untuk kedua dataset, meskipun untuk cpDNA, populasi XA juga menunjukkan tingkat divergensi genetik yang relatif tinggi (Tabel S1 dan S2 ). Memang, nilai F ST rata-rata antara populasi yang termasuk dalam kelompok yang berbeda jauh lebih tinggi ketika dipisahkan oleh bendungan, yaitu, JF (= wilayah SW) versus wilayah NW, daripada ketika dipisahkan oleh jalan bebas hambatan (sisa nilai berpasangan; Tabel S3 dan S4 ); untuk nrDNA, F ST-bendungan[2022] = 0,0113 ± 0,0013 dan F ST-bendungan[2016] = 0,0085 ± 0,0010 versus F ST-expwy[2022] = 0,0075 ± 0,0017 dan F ST-expwy[2016] = 0,0062 ± 0,0020; untuk cpDNA, F ST-dam[2022] = 0,0170 ± 0,0008 dan F ST-dam[2016] = 0,0193 ± 0,0010 versus F ST-expwy[2022] = 0,0110 ± 0,0077 dan F ST-expwy[2016] = 0,0122 ± 0,0030. Jadi, seperti yang juga terjadi di antara populasi individu, nilai F ST di antara kelompok populasi meningkat selama periode 2016–2022 hanya untuk nrITS. Akan tetapi, untuk kumpulan data nrDNA dan cpDNA, tidak ada perbedaan statistik ( p > 0,05) dalam estimasi F ST berpasangan antara tahun 2016 dan 2022.
Berdasarkan pendekatan Δ K , K = 3 adalah jumlah klaster genetik yang paling mungkin setelah menjalankan perangkat lunak STRUKTUR (Gambar S1a,b ), baik untuk kumpulan data nrDNA tahun 2016 maupun 2022. Dalam skema pengelompokan K = 3, satu klaster mencakup populasi BW, YJ, EX, dan YG, yang mewakili kelompok populasi NW (Gambar 4a,b ). Kelompok populasi SE (QX dan WZ) membentuk klaster lain, sementara populasi XA menunjukkan dua komponen genetik (NW dan SE, dengan NW yang dengan cepat mengurangi bobotnya dari tahun 2016 hingga 2022). Populasi JF merupakan klaster ketiga.
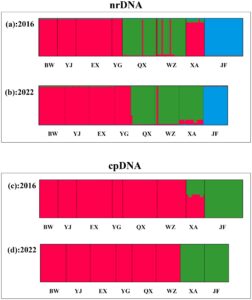
Pengelompokan Bayesian menggunakan sekuens cpDNA agak berbeda dibandingkan dengan nrDNA (Gambar 4c,d ), karena hanya ada dua klaster genetik ( K = 2) (Gambar S1c,d ), yang menunjukkan bahwa dam bertindak sebagai penghalang genetik utama di kedua kumpulan data. Klaster pertama mencakup semua populasi kecuali JF (yaitu, kelompok SW) dan sebagian XA (yaitu, kelompok S) untuk kumpulan data 2016, sementara 6 tahun kemudian populasi XA masuk dalam klaster kedua.
Hasil AMOVA menunjukkan bahwa sebagian besar variasi genetik ITS berada di antara keempat wilayah (Tabel 4 ), meskipun komponen ini menurun dari 71% (2016) menjadi 62% (2022), dan komponen dalam populasi meningkat dari 9% (2016) menjadi 16% (2022). Tidak seperti kumpulan data nrDNA, variasi genetik cpDNA yang berada di antara wilayah meningkat dari tahun 2016 (72%) menjadi tahun 2022 (90%), sedangkan komponen dalam populasi menurun dari 10% (2016) menjadi 4% (2022).
| Derajat kebebasan | Jumlah kuadrat | Perkiraan variasi | Variasi (%) | |
|---|---|---|---|---|
| DIA | ||||
| Tahun 2016 | ||||
| Diantara wilayah | 3 | 371.793 | 2.617 | 71 |
| Diantara pop. | 4 | 56.858 | 0.751 | 20 |
| Dalam pops. | 168 | 55.099 | 0.328 | 9 |
| Total | 175 | 483.750 | 3.66 | 100 |
| Tahun 2022 | ||||
| Diantara wilayah | 3 | 218.566 | 2.310 | 62 |
| Diantara pop. | 4 | 50.063 | 0.832 | 22 |
| Dalam pops. | 109 | 65.133 | 0,598 | 16 |
| Total | 116 | 333.752 | 3.739 | 100 |
| DNA-Cp | ||||
| Tahun 2016 | ||||
| Diantara wilayah | 3 | 1313.175 | 9.308 | 72 |
| Diantara pop. | 4 | 192.434 | 2.412 | 18 |
| Dalam pops. | 168 | 210.613 | 1.254 | 10 |
| Total | 175 | 1716.222 | 12.974 | 100 |
| Tahun 2022 | ||||
| Diantara wilayah | 3 | 998.455 | 12.328 | 90 |
| Diantara pop. | 4 | 52.041 | 0,868 | 6 |
| Dalam pops. | 109 | 62.529 | 0,574 tahun | 4 |
| Total | 116 | 1113.026 | 13.770 | 100 |
3.4 Hambatan Geografis, Isolasi Berdasarkan Jarak, Analisis Distribusi Ketidaksesuaian, dan Deteksi Netralitas
Dengan menghitung perbedaan maksimum Monmonier menggunakan program BARRIER, kami mendeteksi tiga penghalang geografis menggunakan nrDNA dan cpDNA secara terpisah, meskipun penghalang ini sedikit berbeda dalam posisi dan signifikansi dengan penanda yang digunakan dan tahun yang dinilai. Untuk sampel tahun 2016, penghalang pertama menggunakan nrDNA (didukung oleh 99% matriks F ST ) memisahkan QX dari WZ dan XA, sehingga sebagian sesuai dengan Expressway 2 (Gambar 5a ). Penghalang kedua (dukungan 66%) memisahkan kelompok NW dari populasi lain, sehingga cocok dengan Waduk Daguangba dan Expressway 1. Akhirnya, penghalang ketiga (dukungan 33%) ditemukan antara populasi XA dan WZ (Gambar 5a ). Untuk sampel tahun 2022, penghalang pertama (dukungan 99%) memisahkan populasi JF dari sisanya (yaitu, Waduk Daguangba plus bagian dari Expressway 1) (Gambar 5b ). Bagian dari penghalang kedua (66% mendukung) dan ketiga (33% mendukung) juga dapat ditetapkan pada beberapa bagian jalan tol bebas hambatan 1 dan 2, masing-masing (Gambar 5b ).
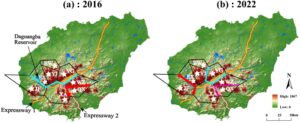
Perhitungan penghalang berdasarkan matriks sekuens cpDNA menunjukkan pola yang serupa dengan yang diperoleh dengan nrDNA. Untuk sampel tahun 2016, satu-satunya perbedaan adalah lokasi penghalang ketiga, yang untuk cpDNA memisahkan YJ dari BW dan YG (Gambar 6a ). Untuk sampel tahun 2022, penghalang pertama berubah menjadi reservoir tetapi juga mencakup beberapa bagian jalan tol 1 dan 2 (Gambar 6b ). Penghalang kedua sesuai dengan bagian Jalan Tol 1 yang terletak di antara YG dan WZ, sedangkan penghalang ketiga terletak di antara YJ dan YG (Gambar 6b ).
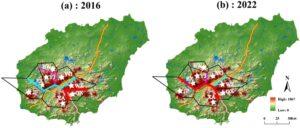
Uji Mantel menunjukkan korelasi positif yang signifikan antara jarak genetik berpasangan dan jarak geografis untuk nrDNA dan cpDNA selama dua periode penelitian (Gambar S2 ). Hasil uji Tajima’s D dan uji Fu’s F s disajikan dalam Tabel 2 dengan nilai- p terkait . Nilai untuk D dan F s positif untuk urutan nrDNA 2016 ( D = 0,80474, p > 0,10; F s = 1,811), urutan cpDNA 2016 ( D = 1,38922, p > 0,10; F s = 12,439), dan urutan cpDNA 2022 ( D = 2,2589, p < 0,05; F s = 11,3), tetapi negatif untuk urutan nrDNA 2022 ( D = −1,18345, p > 0,10; F s = −3,937) (Tabel 2 ). Hasil ini menunjukkan bahwa urutan kami (dengan satu-satunya pengecualian dari urutan cpDNA 2022) sesuai dengan hipotesis nol tentang ukuran populasi konstan dan evolusi netral. Analisis ketidaksesuaian hierarkis menunjukkan distribusi perbedaan untuk semua populasi (Gambar S3 ), yang darinya hipotesis perluasan populasi demografis dapat ditolak.
4 Diskusi
4.1 Dampak Cepat Hambatan Antropogenik terhadap Pola Filogeografi
Evaluasi dampak fragmentasi habitat pada spesies hewan dan tumbuhan secara tradisional ditangani dengan membandingkan aliran gen, diferensiasi genetik, dan struktur genetik populasi di habitat terfragmentasi dan tidak terfragmentasi secara bersamaan (Chung, Nason, et al. 2014 ; Gao et al. 2015 ; Schlaepfer et al. 2018 ). Misalnya, Su et al. ( 2003 ) membandingkan diferensiasi genetik populasi yang dipisahkan oleh Tembok Besar China dengan populasi yang tidak mengalami pemisahan fisik yang besar ini selama enam tanaman dan menemukan perbedaan yang dapat diatribusikan pada keberadaan tembok selama 600 tahun. Dalam contoh kedua, tingkat divergensi genetik pada dua udang karang ( Faxonius validus dan F. erichsonianus ) secara signifikan lebih tinggi di sungai yang dibendung selama 36–104 tahun daripada di sungai yang tidak dibendung di Alabama, Amerika Serikat (Barnett et al. 2019 ). Perbandingan langsung (yaitu, membandingkan populasi yang benar-benar mengalami gangguan tersebut sebelum dan sesudah peristiwa fragmentasi) mungkin tidak tersedia dalam literatur karena diasumsikan bahwa waktu yang berlalu sejak fragmentasi seharusnya sangat lama, karena cukup banyak generasi yang telah berlalu untuk mengamati perubahan genetik. Dalam meta-analisis mereka, Schlaepfer et al. ( 2018 ) menemukan bahwa efek umumnya hanya dapat diamati setelah 50 tahun, meskipun beberapa pengecualian dapat berlaku, seperti kasus di mana hanya 1–5 generasi yang telah berlalu.
Di sini kami telah mengukur efek fragmentasi konstruksi antropogenik dengan menggunakan populasi tanaman yang sama ( P. heterotricha ) dua kali dalam 6 tahun (yaitu, setidaknya dua generasi tanaman studi akan berlalu). Kami telah mendeteksi struktur genetik yang signifikan di seluruh populasi (Gambar 3 dan 4 ), dan AMOVA menunjukkan tingkat variasi genetik yang tinggi antara wilayah (Tabel 4 ), yang menunjukkan diferensiasi genetik yang signifikan dan aliran gen yang terbatas di antara populasi dan wilayah. Baik pohon filogenetik dan hasil STRUKTUR menunjukkan bahwa ada tiga garis keturunan genetik yang berbeda, yang sesuai dengan tiga klade; yaitu, klade barat laut (wilayah NW), klade barat daya (wilayah SW), dan klade tenggara (wilayah SE) (Gambar 3 dan 4 ). Seperti yang diharapkan untuk spesies yang menghuni daerah pegunungan terjal (dan di mana beberapa sungai dibendung sejak akhir abad ke-20), struktur genetik yang dapat dideteksi sudah ada sebelum pembangunan jalan bebas hambatan (2016). Walaupun dampaknya belum begitu besar (mungkin karena waktu yang berlalu belum cukup), kami telah mampu mendeteksi beberapa perubahan pada struktur genetik P. heterotricha hanya beberapa tahun setelah dua jalan tol tersebut selesai dibangun.
Tiga klade yang diidentifikasi untuk P. heterotricha kemungkinan mendahului pembentukan penghalang antropogenik Waduk Daguangba dan Jalan Tol 1 dan 2, karena jangkauan spesies ini, yang terdapat di pegunungan pada ketinggian yang relatif tinggi, secara alami terfragmentasi oleh lembah-lembah sungai. Seperti yang dapat diamati pada Gambar 2 (dan Gambar 5 dan 6 ), Jalan Tol 1 telah dibangun di sepanjang Sungai Changhua (yang lembahnya, di beberapa bagian, selebar 3 km) yang melintasi sistem pegunungan tengah-selatan Hainan dalam arah barat daya–timur laut, dan kemudian sungai berbelok ke barat laut, di mana ia dibendung oleh Waduk Daguangba. Jalan Tol 2 telah dibangun dengan memanfaatkan lembah Sungai Tongshi meskipun jauh lebih sempit. Namun, seperti yang ditunjukkan hasil kami, konstruksi antropogenik akan mengintensifkan efek isolasi Sungai Changhua dan lembah terkaitnya, yang telah menjadi penghalang geografis sejak lama (Xiao et al. 2012 ). Selain itu, jalan konvensional dibangun di lembah-lembah sungai ini jauh lebih awal; misalnya, jalan G224, yang selesai dibangun pada tahun 1954, membentang hampir di rute yang sama dengan kedua jalan bebas hambatan tersebut.
Perubahan signifikansi penghalang sebelum dan setelah pembangunan jalan dalam analisis BARRIER (Gambar 5 dan 6 ) menunjukkan bahwa isolasi geografis yang disebabkan oleh konstruksi manusia merupakan kunci untuk memahami pola filogeografis P. heterotricha saat ini . Sebagian besar penghalang pertama untuk nrDNA dan cpDNA untuk tahun 2016 sesuai dengan pemisahan antara populasi QX dan WZ, yang tidak sesuai dengan konstruksi antropogenik mana pun. Sebaliknya, untuk tahun 2022, penghalang pertama bertepatan dengan bendungan dan jalan bebas hambatan untuk kedua urutan. Sinyal lain yang tak terbantahkan yang menunjukkan peran penting hambatan antropogenik pada struktur genetik P. heterotricha adalah perubahan tajam dalam afinitas genetik populasi XA, baik untuk nrDNA dan cpDNA (Gambar 4 ), yang mungkin disebabkan oleh isolasi Expressway 1 tetapi terutama Expressway 2. Meskipun sulit untuk membedakan kontribusi relatif dari lembah dan sungai di satu sisi, dan jalan bebas hambatan di sisi lain, perubahan yang dijelaskan di atas dalam pola genetik dari tahun 2016 hingga 2022 menunjukkan bahwa kedua jalan bebas hambatan tersebut mungkin berkontribusi untuk membentuk pola filogeografis P. heterotricha endemik pulau tepat setelah 6 tahun berada di tempatnya, kemungkinan sesuai dengan dua generasi tanaman. Ketidaksinambungan geografis, termasuk gangguan antropogenik, merupakan faktor penting dalam diferensiasi populasi dengan melemahkan atau menghalangi aliran gen pada banyak spesies tumbuhan (Slatkin 1985 ; Su et al. 2003 ; Kartzinel et al. 2013 ; Chung, López-Pujol, et al. 2014 ; Almeida-Rocha et al. 2020 ). Efek ini bahkan lebih besar untuk tumbuhan herba kecil seperti P. heterotricha , dengan potensi penyebaran yang buruk yang terkait dengan benih kecil yang sebagian besar disebarkan oleh air.
4.2 Dampak Hambatan Antropogenik terhadap Struktur Genetik
Meskipun klade P. heterotricha yang berbeda memiliki dinamika populasi yang stabil, seperti yang terungkap oleh analisis distribusi ketidakcocokan (Gambar S3 ), tanaman tersebut akan mengalami beberapa efek yang dapat dideteksi pada struktur genetiknya, yang mungkin disebabkan oleh pertukaran gen yang berkurang. Khususnya, pembagian ribotipe di antara tiga kelompok populasi yang dipisahkan oleh jaringan jalan bebas hambatan hampir menghilang. Meskipun tidak ada perbedaan statistik dalam F berpasangan ST berpasangan antara tahun 2016 dan 2022, peningkatan derajat baik pada tingkat populasi ( F ST[2016] = 0,0045, F ST[2022] = 0,0054; Tabel S1 ) dan pada tingkat wilayah (Tabel S3 ) ditemukan dari tahun 2016 hingga 2022. Sebaliknya, F ST tetap tidak berubah seiring waktu dengan cpDNA pada tingkat populasi (0,0098 pada tahun 2016 vs. 0,0097 pada tahun 2022; Tabel S2 ) dan bahkan meningkat pada tingkat kelompok ( F ST hanya menurun antara wilayah NW dan SE, yang dapat dikaitkan dengan pembangunan Jalan Tol 1; Tabel S4 ). Ketidaksesuaian antara nrDNA dan cpDNA ini dapat berasal dari kedua cara pewarisan yang berbeda (nrDNA diwariskan secara biparental, cpDNA hanya diwariskan secara maternal) dan dari fakta bahwa laju migrasi serbuk sari seringkali jauh lebih tinggi daripada laju migrasi benih (Ennos 1994 ; Petit et al. 2005 ). Dengan demikian, seseorang dapat memperkirakan bahwa cpDNA yang diwariskan secara maternal (yang hanya ditularkan melalui benih) akan jauh kurang sensitif terhadap efek fragmentasi, khususnya jika kita memperhitungkan bahwa hambatan antropogenik (terutama bendungan) akan memengaruhi aliran serbuk sari pada tingkat yang jauh lebih besar daripada aliran benih (lihat di bawah).
Diferensiasi genetik lebih tinggi antara populasi yang dipisahkan oleh bendungan daripada antara populasi yang dipisahkan oleh jalan bebas hambatan (Tabel S3 dan S4 ); yaitu, Waduk Daguangba menunjukkan efek penghalang yang lebih jelas pada aliran gen. Dua faktor dapat menjelaskan pola yang diamati seperti itu. Pertama, waduk dibangun pada tahun 1994; oleh karena itu, periode isolasi hampir 30 tahun berpotensi menghasilkan lebih banyak konsekuensi genetik daripada jalan bebas hambatan yang baru selesai dibangun, dengan periode efek 6 tahun atau dua generasi P. heterotricha . Kedua, bendungan memiliki waduk air sepanjang sekitar 6 km dan permukaan air 100 km 2. Dengan demikian, bendungan mungkin membawa lebih banyak efek penghalang daripada jalan bebas hambatan, yang biasanya hanya selebar 20–30 m. Biji P. heterotricha berukuran kecil dan sebagian besar dapat disebarkan oleh tetesan air hujan atau aliran air, sementara penyerbukan spesies ini membutuhkan serangga berukuran kecil seperti Amegilla leptocoma dan A. yunnanensis (Ling et al. 2017a ). Mekanisme penyebaran biji mungkin dipengaruhi oleh Waduk Daguangba, karena bendungan mampu membatasi hidrokori (Andersson et al. 2000 ). Namun, dampaknya akan jauh lebih luas terkait penyebaran serbuk sari, karena jarak terbang serangga kecil ini hampir tidak cukup untuk menghubungkan kedua tepi waduk (terpisah hingga 4 km).
Meskipun dampaknya lebih kecil daripada waduk, dampak jalan bebas hambatan masih terlihat untuk P. heterotricha . Selain penghalang fisik yang ditimbulkan oleh dua jalan berkapasitas tinggi, pertumbuhan lalu lintas yang cepat (terutama setelah berakhirnya pandemi COVID-19)—dengan lebih banyak kebisingan dan akumulasi polutan—akan meningkatkan tingkat gangguan. Lalu lintas yang bergerak cepat dan perubahan kondisi angin yang terkait dengan jalan bebas hambatan mungkin memengaruhi pergerakan penyerbuk P. heterotricha , sehingga mengurangi aliran serbuk sari. Dampak negatif jalan terhadap pergerakan (dan bahkan kelangsungan hidup) penyerbuk sering terdeteksi (Stephens et al. 2000 ; Bhattacharya et al. 2003 ; Baxter-Gilbert et al. 2015 ; Fitch and Vaidya 2021 ; Dániel-Ferreira et al. 2022 ). Meskipun pembangunan jalan dapat mengubah kondisi hidrologi (dengan memengaruhi jalur aliran alami dan kualitas air, misalnya, Buchanan et al. 2013 ), kami belum dapat menemukan contoh dalam literatur tentang jalan yang memengaruhi hidrokori.
Sebagai kesimpulan, kami memberikan bukti bahwa efek penghalang yang disebabkan oleh konstruksi antropogenik (seperti waduk dan jalan bebas hambatan) dapat menghasilkan perubahan dalam struktur genetik dan pola filogeografis hanya dalam dua generasi tanaman ini. Efek disruptif yang melibatkan fragmentasi habitat ini dapat menimbulkan ancaman utama terhadap regenerasi populasi, keanekaragaman genetik, dan mengubah proses evolusi spesies, terutama pada tanaman endemik yang terancam punah dan berumur pendek. Untuk mengurangi tekanan negatif tersebut, kami sarankan untuk membangun koridor ekologi guna meningkatkan aliran gen antara populasi yang dipisahkan oleh penghalang antropogenik ini. Ini dapat mencakup terowongan jalan dan hutan layang, yang memfasilitasi penyebaran tanaman. Koridor ekologi tersebut juga dapat meningkatkan integritas vegetasi dan kesinambungan habitat antara kedua sisi penghalang ini dan dengan demikian membantu persistensi jangka panjang P. heterotricha endemik pulau dan spesies langka dan terancam punah lainnya.