
Abstrak
Interaksi frugivori merupakan proses ekologi penting untuk regenerasi hutan tropis, yang memastikan ketahanan ekosistem setelah gangguan. Namun, sedikit yang diketahui tentang bagaimana interaksi frugivori dibentuk oleh gangguan antropogenik, terutama di Amazonia – salah satu ekosistem dengan keanekaragaman hayati tertinggi di Bumi. Kami menyelidiki bagaimana penebangan selektif dan kebakaran hutan memengaruhi interaksi frugivori arboreal dan terestrial di hutan Amazon. Kami fokus pada empat kelas hutan: Tidak terganggu, Ditebang, Ditebang-dan-17-tahun-terbakar (terbakar 17 tahun sebelum pengambilan sampel) dan Ditebang-dan-3-tahun-terbakar (terbakar tiga tahun sebelum pengambilan sampel). Kami mencatat 4670 interaksi frugivori di tingkat komunitas, dalam upaya pengambilan sampel selama 31.484 jam. Hutan yang tidak terganggu mempertahankan jumlah spesies dan interaksi yang jauh lebih tinggi jika dibandingkan dengan hutan yang Ditebang-dan-17-tahun-terbakar, dan jumlah yang sama dengan yang Ditebang dan Ditebang-dan-3-tahun-terbakar. Penebangan selektif dan kebakaran hutan tidak mengubah secara signifikan sifat struktural jaringan pemakan buah, yang sangat modular, cukup terspesialisasi, kurang terhubung dan tidak bersarang. Mengenai komposisi komunitas, kami mendeteksi keragaman β yang tinggi dari spesies tumbuhan, spesies pemakan buah, dan interaksinya antara semua area studi, serta di dalam dan antara kelas hutan, terutama didorong oleh pergantian spesies. Hutan yang ditebang dan dibakar selama 17 tahun menjadi tuan rumah komposisi interaksi yang paling unik dibandingkan dengan hutan yang tidak terganggu. Studi kami memberikan bukti baru bahwa gangguan antropogenik, khususnya penebangan selektif dan kebakaran hutan, berdampak negatif pada interaksi pemakan buah di hutan Amazon. Efek ini dapat bertahan selama bertahun-tahun setelah peristiwa gangguan awal dan dapat diperburuk karena peningkatan kebakaran hutan yang diprediksi didorong oleh perubahan iklim.
Perkenalan
Di Amazon, 38% habitat hutan telah terdampak oleh berbagai jenis gangguan antropogenik, yang sering kali beraksi secara sinergis (Díaz et al. 2019 , Lapola et al. 2023 ). Misalnya, antara tahun 2001 dan 2018, penebangan selektif telah memengaruhi 120.000 km 2 di wilayah tersebut (Lapola et al. 2023 ). Hutan yang ditebang secara selektif lebih panas dan kering karena lebih banyaknya pembukaan lahan, sehingga membuatnya lebih rentan terhadap kebakaran hutan (Uhl dan Kauffman 1990 , Cochrane 2003 ). Peningkatan frekuensi, intensitas, dan lamanya kekeringan ekstrem di seluruh wilayah juga mengakibatkan lebih banyak hutan yang mudah terbakar (Cochrane 2003 , Nepstad et al. 2004 , Jiménez-Muñoz et al. 2016 ). Sebagai konsekuensinya, kebakaran hutan diperkirakan akan menyebar sangat luas pada abad ke-21 (Brando et al. 2020 , Lapola et al. 2023 ), dengan beberapa kebakaran besar, yaitu kebakaran yang mempengaruhi area ≥ 10.000 ha, telah dilaporkan (Withey et al. 2018 ).
Mengingat bahwa hutan Amazon tidak berevolusi dalam kondisi gangguan kebakaran alam yang teratur (Uhl dan Kauffman 1990 ), peristiwa seperti itu menyebabkan kematian pohon yang tinggi (Berenguer et al. 2021 ). Komunitas tumbuhan pasca-kebakaran sering didominasi oleh spesies pionir dengan kepadatan kayu rendah dan biji kecil (Barlow et al. 2003 , Barlow dan Peres 2004 , Hawes et al. 2020 ). Spesies pionir ini juga merupakan koloni umum hutan yang baru-baru ini mengalami penebangan selektif (Costantini et al. 2016 ). Dalam jangka panjang, pergeseran komposisi spesies ini dapat membahayakan ketersediaan buah-buahan untuk fauna hutan, yang berpotensi menyebabkan penurunan tingkat kunjungan frugivora (Moegenburg dan Levey 2003 , Brando et al. 2024 ). Perubahan ketersediaan buah seperti itu juga dapat mengubah komposisi komunitas frugivora, terutama yang memengaruhi spesies frugivora yang ketat. Memang, komunitas burung pemakan buah di hutan tropis yang terganggu terbukti kurang beragam dan berbeda dalam komposisi spesies jika dibandingkan dengan hutan yang tidak terganggu (Moura et al. 2013 , Bregman et al. 2016 ). Pergeseran seperti itu dalam komunitas tumbuhan dan pemakan buah kemungkinan akan mengganggu proses ekologi penting seperti penyebaran benih dan pemangsaan benih, yang pada gilirannya akan memengaruhi regenerasi alami dan, akhirnya, dinamika komunitas pasca-gangguan (Valiente-Banuet et al. 2015 , Neuschulz et al. 2016 , Rogers et al. 2021 ).
Meskipun ada kemajuan dalam pemahaman kita tentang dampak gangguan antropogenik pada komunitas tumbuhan dan pemakan buah, tidak ada penelitian yang membahas dampak ini pada interaksi pemakan buah di tingkat komunitas di hutan Amazon (Ballarin et al. 2024 ). Hingga saat ini, studi di wilayah tersebut terutama berfokus pada penggambaran interaksi spesies tunggal, seperti tapir (Fragoso dan Huffman 2000 , Tobler et al. 2010 , Paolucci et al. 2019 ), atau kelompok spesifik seperti macaw (Baños-Villalba et al. 2017 ), kura-kura dan ikan (Guzmán dan Stevenson 2008 , Weiss et al. 2016 ), tumbuhan kunci (Stevenson et al. 2015 ), dan/atau serikat vertebrata diurnal berbadan sedang hingga besar (Hawes dan Peres 2014 ). Penelitian yang melibatkan seluruh komunitas pemakan buah, termasuk pemakan buah arboreal dan terestrial, menimbulkan tantangan yang signifikan, mengingat luasnya jangkauan vertebrata pemakan buah dan jenis buah morfologi yang mereka eksploitasi (Hawes dan Peres 2014 , Todeschini et al. 2020 , Scabin dan Peres 2021 ).
Jaringan ekologi sering digunakan untuk memahami bagaimana interaksi pemakan buah terstruktur (Delmas et al. 2019 ). Penggunaan jaringan juga memberikan wawasan berharga tentang bagaimana gangguan membentuk kembali interaksi ekologi (Tylianakis dan Morris 2017 , Emer dan Timóteo 2020 ). Setelah gangguan antropogenik, beberapa sifat struktural jaringan diperkirakan akan berubah (Menke et al. 2012 , de Assis Bomfim et al. 2018 ). Misalnya, konektivitas jaringan – dipahami sebagai proporsi interaksi yang terwujud relatif terhadap semua kemungkinan kombinasi spesies berpasangan dalam sistem (Dunne et al. 2002 ) – diperkirakan akan meningkat setelah gangguan antropogenik, karena jumlah spesies biasanya menurun dan oleh karena itu kemungkinan interaksi yang terwujud di antara yang tersisa cenderung meningkat (Heleno et al. 2012 , Menke et al. 2012 ). Keterkaitan jaringan – yang menunjukkan sejauh mana interaksi spesies spesialis merupakan bagian dari interaksi yang melibatkan spesies generalis, sementara interaksi antara spesies generalis tidak umum (Bascompte et al. 2003 , Almeida-Neto et al. 2008 ) juga diprediksi meningkat, karena spesies generalis cenderung menjadi sangat berlimpah di ekosistem yang terganggu (Emer et al. 2020 ). Modularitas – ukuran apakah kelompok spesies memiliki interaksi yang lebih kuat di antara mereka sendiri dibandingkan dengan spesies dari kelompok lain (Olesen et al. 2007 ) – diperkirakan akan menurun setelah peristiwa gangguan, karena interaksi spesialis cenderung menghilang (Schleuning et al. 2014 ). Terakhir, spesialisasi jaringan – yang mencerminkan bagaimana spesies secara eksklusif berinteraksi satu sama lain (Vázquez dan Aizen 2004 , Blüthgen et al. 2006 ) – juga diperkirakan akan menurun karena interaksi generalis cenderung menjadi lebih umum di hutan yang terganggu (Menke et al. 2012 , Palacio et al. 2016 ). Selain analisis jaringan, dampak gangguan antropogenik juga dapat dinilai dengan memeriksa perbedaan spesies di antara berbagai area – β-keanekaragaman (Poisot et al. 2012 , Burkle et al. 2022 ). Misalnya, penggantian spesies asli atau perubahan interaksi spesies dapat mengakibatkan pergantian ekologis (misalnya β-pergantian), termasuk kasus-kasus di mana spesies yang tersisa adalah subset bersarang dari spesies asli (β-nestedness, Baselga 2010 ).
Di sini, kami menggunakan kedua sifat struktur jaringan dan metrik β-diversity untuk mengukur dampak penebangan selektif dan kebakaran hutan pada interaksi pemakan buah di hutan Amazon. Desain pengambilan sampel kami berfokus pada tingkat komunitas, termasuk pemakan buah arboreal dan terestrial, untuk mengambil sampel seluruh kelompok fungsional pemakan buah vertebrata. Kami mengambil sampel interaksi pemakan buah di empat kelas hutan: 1) hutan yang tidak terganggu, 2) hutan yang ditebang secara selektif, 3) hutan yang ditebang secara selektif yang terbakar setidaknya 17 tahun sebelum pengambilan sampel, dan 4) hutan yang ditebang secara selektif yang terbakar tiga tahun sebelum pengambilan sampel. Untuk kemudahan referensi, kelas-kelas hutan ini masing-masing disebut sebagai Tidak Terganggu, Ditebang, Ditebang-dan-Terbakar-17-tahun dan Ditebang-dan-Terbakar-3-tahun. Secara khusus, kami menyelidiki bagaimana penebangan selektif dan kebakaran hutan memengaruhi:
1) jumlah tumbuhan dan spesies pemakan buah, serta interaksinya;
2) sifat-sifat struktur jaringan pemakan buah (yaitu konektivitas, keterkaitan, modularitas dan spesialisasi);
3) keanekaragaman β tumbuhan, pemakan buah, dan interaksinya.
Kami berhipotesis bahwa akan ada penurunan jumlah tumbuhan dan spesies pemakan buah setelah penebangan selektif dan kebakaran hutan (Moura et al. 2013 , Berenguer et al. 2021 ), dengan implikasi langsung untuk interaksi pemakan buah (Albert et al. 2020 ), mengurangi frekuensinya dan mengubah asosiasi spesies berpasangan. Kami memperkirakan bahwa dampak akan lebih kuat di hutan yang ditebang secara selektif dan dibakar baru-baru ini, di mana kami memperkirakan lebih sedikit spesies dan interaksi dibandingkan dengan hutan yang tidak terganggu karena kebakaran baru-baru ini. Sebaliknya, kami memperkirakan bahwa hutan yang ditebang secara selektif dan hutan yang ditebang secara selektif dan dibakar hampir dua dekade lalu akan menampung lebih banyak spesies dan interaksi daripada hutan yang terbakar baru-baru ini, mengingat periode pemulihan pasca kebakaran yang panjang. Dalam kasus hutan yang ditebang, kami berharap bahwa hutan tersebut akan menjadi yang paling kecil dampaknya, mengingat tidak adanya peristiwa kebakaran. Lebih jauh, kami berhipotesis bahwa interaksi yang tersisa di hutan yang ditebang secara selektif dan dibakar baru-baru ini akan menghasilkan struktur jaringan yang lebih terhubung, lebih bersarang, kurang modular, dan kurang terspesialisasi dibandingkan dengan hutan yang tidak terganggu (Sebastián-González et al. 2015 , Tylianakis dan Morris 2017 ). Ini karena spesies generalis diperkirakan akan menang di area ini, berkontribusi secara tidak proporsional terhadap interaksi berpasangan, sambil mengurangi jumlah spesies spesialis (Burivalova et al. 2014 , Hawes et al. 2020 ). Akhirnya, kami memprediksi nilai β-diversitas yang tinggi untuk spesies dan interaksi antara area studi dan antara kelas hutan. Nilai tertinggi diperkirakan antara hutan yang ditebang secara selektif dan dibakar baru-baru ini dan hutan yang tidak terganggu, karena perbedaan komposisi spesies dan interaksi yang dihasilkan dari peristiwa kebakaran hutan baru-baru ini. Kebakaran ini mendorong pergantian spesies dan interaksi, yaitu pergantian β (lihat Burkle et al. 2022 untuk interaksi penyerbukan). Mengenai hutan yang ditebang secara selektif dan ditebang secara selektif dan dibakar hampir dua dekade lalu, kami memperkirakan hutan ini lebih mirip dengan hutan yang tidak terganggu mengingat waktu pemulihan hampir dua dekade. Periode pemulihan yang diperpanjang ini memfasilitasi keberadaan tidak hanya spesies umum tetapi juga spesies yang terkait dengan hutan yang tidak terganggu.
Bahan dan metode
Daerah studi
Studi ini dilakukan di Amazon Brasil bagian timur, di munisipalitas Belterra, Mojuí dos Campos dan Santarém (Informasi pendukung). Wilayah ini terdiri dari mosaik penggunaan lahan, termasuk hutan primer yang tidak terganggu, hutan primer yang terdegradasi, hutan sekunder dengan umur yang bervariasi, lahan pertanian dan padang rumput (Gardner et al. 2013 ). Penebangan selektif dan kebakaran hutan merupakan beberapa ancaman utama bagi wilayah hutan yang tersisa (Lapola et al. 2023 ). Selama tahun 2015 dan 2016, kekeringan ekstrem, yang disebabkan oleh peristiwa El Niño yang kuat, menyebabkan kebakaran yang meluas ke lebih dari satu juta hektar hutan di wilayah studi. Hutan yang ditebang secara selektif dan dibakar baru-baru ini (Hutan yang ditebang dan dibakar selama 3 tahun) dalam studi ini adalah hutan yang terbakar selama peristiwa El Niño ini (Withey et al. 2018 ).
Desain pengambilan sampel
Kami mempelajari 17 area di empat kelas hutan: 1) Hutan tak terganggu – yaitu hutan primer tanpa bukti gangguan antropogenik (n = 5), 2) Hutan tebangan – yaitu hutan primer tebangan selektif (n = 4), 3) Hutan tebangan dan terbakar 17 tahun – yaitu hutan primer tebangan selektif yang terbakar sedikitnya 17 tahun sebelum pengambilan sampel (n = 4), 4) Hutan tebangan dan terbakar 3 tahun – yaitu hutan primer tebangan selektif yang terbakar tiga tahun sebelum pengambilan sampel (n = 4, Informasi pendukung). Jika ada, penebangan selektif terjadi sedikitnya 30 tahun sebelum penelitian. Jenis dan waktu gangguan antropogenik yang memengaruhi setiap area penelitian ditentukan melalui penilaian lapangan dan inspeksi visual citra satelit dari tahun 1988 hingga 2010 (Gardner et al. 2013 ). Di setiap area studi, kami memasang transek tunggal sepanjang 1 km, memastikan bahwa setiap transek dipisahkan dari yang lain setidaknya 1,5 km untuk meminimalkan autokorelasi spasial (Informasi pendukung).
Pengambilan sampel interaksi frugivori
Dari Februari 2019 hingga Maret 2020, kami mencatat interaksi frugivori antara tanaman berbuah berdaging dan burung dan mamalia frugivora (kecuali kelelawar). Interaksi frugivori dicatat pada tingkat komunitas tanpa berfokus pada kelompok taksonomi tanaman atau spesies frugivora tertentu. Perilaku makan berikut dianggap sebagai interaksi frugivori: menelan biji – frugivora menelan seluruh biji; konsumsi pulp – frugivora memakan buah sepotong-sepotong tetapi tidak menelan biji; biji terbawa – frugivora mengambil buah dari tanaman alih-alih segera menelannya; dan predasi biji – frugivora merusak biji (Jordano dan Schupp 2000 ). Untuk setiap interaksi, kami mencatat tanaman dan spesies frugivora, serta jumlah individu yang berinteraksi dengan buah.
Kunjungan pemakan buah ke masing-masing tanaman diambil sampelnya menggunakan tiga metode yang saling melengkapi untuk memaksimalkan keragaman interaksi yang terekam, termasuk kelompok fungsional arboreal dan terestrial. Untuk menggabungkan ketiga metode tersebut, kami membakukan data dengan mempertimbangkan setiap kunjungan pemakan buah sebagai frekuensi interaksi. Tiga matriks tertimbang (satu untuk setiap metode), yang dibobot berdasarkan frekuensi interaksi, digabungkan dengan menjumlahkan kumpulan data secara langsung (Quintero et al. 2022 ). Meskipun metode tersebut memiliki upaya pengambilan sampel yang berbeda jika dibandingkan satu sama lain, kami memperkirakan upaya pengambilan sampel untuk setiap area studi dan membakukannya di seluruh kelas hutan sejauh yang memungkinkan, untuk memastikan upaya pengambilan sampel tingkat kelas hutan yang serupa (Informasi pendukung).
Tiga metode pengambilan sampel adalah:
1) Sensus berjalan. Interaksi pemakan buah dicatat sepanjang transek sepanjang 1 km antara pukul 06.00 dan 18.00, tanpa survei yang dilakukan selama periode hujan. Catatan pemakan buah diperoleh dengan mengamati tanaman yang berada dalam jarak 25 m dari kedua sisi transek, dengan total 50.000 m² yang diambil sampelnya per transek. Setiap area studi diambil sampelnya dua kali sebulan. Upaya pengambilan sampel untuk sensus berjalan adalah 1.053 jam.
2) Sensus fokus. Sampel fokus dilakukan secara oportunistik selama sensus berjalan. Ketika tanaman dengan buah matang diamati, kami menghentikan sensus berjalan dan memulai pengamatan fokus. Tanaman fokus diamati minimal selama 15 menit. Jika pemakan buah mengunjungi tanaman selama periode ini, waktu pengamatan fokus tambahan selama 30 menit ditambahkan. Upaya pengambilan sampel untuk sensus fokus adalah 503 jam.
3) Kamera penjebakan. Kami membuat 88 titik kamera penjebakan (14 arboreal dan 74 terestrial) menggunakan kamera APEMAN 16MP dengan Infrared Night Vision yang mendeteksi gerakan hingga 20 m jauhnya. Kami memasang 3–6 kamera per area studi pada tanaman selama periode buah matang, melepasnya ketika tidak ada lagi buah yang tersedia. Kamera dipasang menghadap buah untuk memaksimalkan kemungkinan perekaman interaksi (Zhu et al. 2022 ). Kamera arboreal dipasang 5–15 m dari tanah, sedangkan kamera terestrial ditempatkan 30 cm di atas tanah. Kamera aktif 24 jam sehari -1 , merekam video 20 detik ketika dipicu oleh gerakan, dengan interval 20 detik antara perekaman berturut-turut. Video berurutan dari spesies yang sama dianggap independen jika lebih dari satu jam telah berlalu di antara mereka (Rivas-Romero dan Soto-Shoender 2015 ). Ketika lebih dari satu individu direkam dalam satu video, setiap individu dihitung sebagai peristiwa terpisah. Upaya pengambilan sampel dihitung sebagai jumlah jam setiap kamera perangkap aktif, totalnya 29.928 jam.
Spesies tanaman diidentifikasi oleh parabotanis dari Embrapa Amazônia Oriental hingga tingkat taksonomi serendah mungkin. Spesies burung diidentifikasi terutama oleh LCR dengan bantuan ACL dan MAP, dan mamalia kecil diidentifikasi oleh YGR dengan masukan dari peneliti lokal (Ucapan Terima Kasih).
Sifat Struktur Jaringan
Untuk setiap area studi, kami membangun jaringan pemakan buah dari matriks ij , di mana i mewakili spesies tanaman, j mewakili spesies pemakan buah, dan komponen ij menggambarkan hubungan tertimbang di antara mereka, yaitu frekuensi interaksi antara tanaman i dan pemakan buah j . Kami kemudian menghitung beberapa deskriptor jaringan untuk setiap area studi, yaitu: jumlah spesies tanaman dan pemakan buah, ukuran jaringan (yaitu jumlah total spesies), jumlah interaksi berpasangan yang unik, dan jumlah total interaksi. Kami kemudian memperkirakan serangkaian metrik dengan korelasi r < 0,7 di antara mereka sendiri (Informasi pendukung) untuk memaksimalkan informasi pelengkap tentang struktur jaringan pemakan buah, sebagai berikut:
1) Konektivitas ( C ). Proporsi interaksi yang terealisasi dalam kaitannya dengan jumlah total interaksi yang mungkin terjadi (Dunne et al. 2002 );
2) Keterikatan berbobot ( wNODF ). Ukuran kuantitatif keterikatan, yang selanjutnya disebut sebagai keterikatan jaringan, yang mempertimbangkan pengisian matriks yang menurun sesuai dengan frekuensi interaksi. Ini menunjukkan apakah organisasi interaksi pemakan buah diatur oleh inti spesies generalis yang berinteraksi satu sama lain dan juga dengan subset spesies yang lebih spesialis (Ulrich et al. 2009 , Almeida-Neto dan Ulrich 2011 );
3) Modularitas tertimbang. Keberadaan kelompok spesies tumbuhan dan pemakan buah yang berinteraksi kuat satu sama lain, tetapi berinteraksi lebih jarang atau tidak sama sekali dengan spesies dalam kelompok lain (Olesen et al. 2007 ). Kami menggunakan algoritma DIRT_LPA_wb_plu (Beckett 2016 ) untuk memperkirakan tingkat modularitas dan jumlah modul;
4) Spesialisasi ( H2′ ). Mengukur penyimpangan interaksi yang diamati dari yang diharapkan berdasarkan frekuensi interaksi (Blüthgen et al. 2006 ). Semakin unik interaksinya, semakin tinggi nilai H2′ , yang menunjukkan jaringan yang lebih terspesialisasi.
Akhirnya, untuk menguji apakah properti jaringan yang diamati berbeda dari yang diharapkan secara kebetulan, kami membandingkan nilai yang diamati untuk setiap properti jaringan terhadap 100 pengacakan yang dihasilkan menggunakan algoritma Vaznull (Vázquez et al. 2007 ). Algoritma ini mengacak frekuensi interaksi sambil mempertahankan jumlah spesies dan interaksi yang diamati, dengan demikian mempertahankan konektansi konstan (Vázquez et al. 2007 ). Kami kemudian menghitung skor-z masing-masing untuk setiap nilai properti jaringan yang diamati dengan mengurangi rata-rata model nol dan membaginya dengan SD-nya (Dormann et al. 2009 ). Skor-z dan nilai yang diamati ini digunakan sebagai variabel respons dalam model statistik. Metrik jaringan dan model nol diperkirakan menggunakan paket ‘bipartite’ (Dormann et al. 2009 ).
β-keanekaragaman spesies dan interaksi
Kami mengukur β-keanekaragaman spesies tumbuhan (β plants ), spesies frugivora (β frug ), dan interaksi mereka (β int ) pada tiga tingkat untuk menangkap skala perbedaan yang berbeda. Pertama, pada tingkat area studi, kami mengakses perbedaan antara area studi, memperlakukan setiap area studi sebagai replikasi, terlepas dari kelas hutannya. Ini mencerminkan perbedaan di seluruh lokasi geografis. Kedua, pada tingkat kelas intra-hutan, kami mengevaluasi perbedaan dalam kelas hutan yang sama, menggunakan area studi dalam kelas hutan itu untuk direplikasi. Ini menangkap perbedaan antara area studi yang berbagi kelas hutan yang sama. Akhirnya, pada tingkat kelas antar-hutan, kami mengukur perbedaan antara kelas-kelas hutan yang berbeda, memperlakukan setiap kelas hutan sebagai replikasi. Ini mengukur besarnya perbedaan antara kelas-kelas hutan.
Kami kemudian membagi β-diversity menjadi dua komponen: β-turnover dan β-nestedness, keduanya diperkirakan untuk spesies tumbuhan dan pemakan buah, serta untuk interaksi pemakan buah (Baselga 2010 ). Analisis dilakukan dengan menggunakan fungsi beta.multi dari paket ‘betapart’ (Baselga dan Orme 2012 ). Fungsi ini menghitung ketidakmiripan beberapa situs menggunakan ketidakmiripan Sorensen (β SOR ). Fungsi ini selanjutnya membedakan antara kontribusi penggantian spesies dan interaksi (yaitu β-turnover – β SIM ), dihitung menggunakan ketidakmiripan Simpson, dan kehilangan atau perolehan spesies dan interaksi (yaitu β-nestedness – β NES ), diukur menggunakan ketidakmiripan yang dihasilkan oleh nestedness yang diturunkan sebagai perbedaan antara β SOR dan β SIM (Baselga 2010 ). Selain itu, kami menggunakan penskalaan multidimensi non-metrik (NMDS, dua dimensi) untuk secara grafis menggambarkan perbedaan dalam spesies dan komposisi interaksi. Kami menggunakan matriks perbedaan Bray–Curtis untuk mengordinir area studi dan kelas hutan menurut perbedaannya dalam spesies dan komposisi interaksi. Ordinir NMDS dibuat menggunakan fungsi metaMDS dalam paket ‘vegan’ (Oksanen et al. 2015 ).
Analisis statistik
Untuk menguji dampak penebangan selektif dan kebakaran hutan pada properti jaringan, kami menggunakan model linear umum (GLM) dengan fungsi glm dan glm.nb dari paket ‘lme4’ (Bates et al. 2015 ). Kami memeriksa overdispersi dengan paket ‘DHARMa’ (Hartig 2016 ). Model individual dipasang untuk setiap properti struktural jaringan sebagai variabel respons dan kelas hutan sebagai variabel prediktor. Kami memasang distribusi Poisson dengan fungsi tautan log untuk memodelkan data jumlah, termasuk jumlah spesies tumbuhan dan pemakan buah serta ukuran jaringan. Untuk variabel yang menunjukkan overdispersi (yaitu jumlah interaksi berpasangan yang unik dan jumlah total interaksi), kami menerapkan distribusi binomial negatif. Untuk respons kontinu (yaitu model yang diamati dan nol untuk konektivitas, ketersaringan jaringan, modularitas, dan spesialisasi), kami menggunakan distribusi Gaussian. Kami tidak mendeteksi autokorelasi spasial di antara area studi (Informasi Pendukung). Kami menyesuaikan model penuh menggunakan estimasi kemungkinan maksimum dan memilih struktur efek tetap terbaik berdasarkan kriteria informasi Akaike yang dikoreksi untuk ukuran sampel kecil (AICc), menggunakan paket ‘lme4’ (Bates et al. 2015 ). Untuk mengendalikan perbedaan dalam upaya pengambilan sampel di antara kelas hutan, kami menghitung akar kuadrat dari jumlah interaksi dibagi dengan akar kuadrat dari ukuran jaringan, dan memasukkan nilai-nilai ini sebagai kovariat tetap dalam semua model (Schleuning et al. 2012 ). Model dengan delta AICc < 2 dipilih sebagai penjelasan yang masuk akal untuk pola yang diamati (Informasi pendukung). Kami menggunakan bobot bukti Akaike (wAICc) untuk menentukan peringkat kepentingan relatif untuk setiap model (Burnham dan Anderson 2001 ). Kemudian, kami menyajikan koefisien setiap prediktor, varians yang dijelaskan, dan interval kepercayaan (CI) untuk model linier terbaik. Kami memperkirakan CI model menggunakan fungsi confint dari paket ‘lme4’ (Bates et al. 2015 ). Semua analisis dilakukan di R ( www.r-project.org ).
Hasil
Interaksi frugivora
Kami mencatat 4670 interaksi yang melibatkan 991 asosiasi berpasangan unik antara 165 spesies tumbuhan dan 174 spesies pemakan buah, yang menjumlahkan 31.484 jam pengambilan sampel (Informasi pendukung). Delapan puluh enam persen interaksi pemakan buah yang tercatat bersifat arboreal sementara hanya 14% yang tercatat di tanah. Dari kumpulan pemakan buah, 146 diidentifikasi sebagai spesies burung, yang mencakup 3665 interaksi (78,5%), sementara 28 adalah spesies mamalia, yang melakukan 1005 interaksi (21,5%).
Tumbuhan dari famili Melastomataceae, Moraceae, dan Urticaceae terlibat dalam 52% dari total jumlah interaksi (masing-masing 18,3%, 17,8%, dan 15,8%). Spesies tumbuhan yang paling sering terlibat dalam interaksi dengan pemakan buah adalah Coussapoa tessmannii (Urticaceae, 9,7%), diikuti oleh Ficus morphotype 1 (Moraceae, 8,7%), Brosimum acutifolium (Moraceae, 7,1%), dan Miconia pyrifolia (Melastomataceae, 5,3%).
Burung dari famili Ramphastidae (18,2%), Thraupidae (13,7%) dan Pipridae (11%) menyumbang 43% dari total jumlah interaksi. Pemakan buah yang paling umum mencakup tiga spesies burung: Ceratopipra rubrocapilla manakin berkepala merah (8,9%), Ramphastos vitellinus channel-billed toucan (5,1%), Ramphastos tucanus white-throated toucan (3,9%) dan Pteroglossus bitorquatus red-necked aracari (3,9%), dan satu mamalia, Dasyprocta leporina red-rumped agouti (4,3%). Selain itu, kami mencatat spesies yang terancam berinteraksi dengan tanaman, termasuk Lepidothrix iris opal-crowned manakin (diklasifikasikan sebagai Rentan oleh IUCN), yang ditemukan di hutan yang Tidak Terganggu dan Tertebang, serta Aburria cujubi red-throated piping guan (Rentan), Penelope pileata white-crested guan (Rentan), dan Ateles marginatus white-cheeked spider monkey (Terancam), yang diamati di hutan yang Tidak Terganggu, Tertebang, dan Tertebang-dan-Terbakar-3-tahun (IUCN 2024 ). Tidak ada spesies yang terancam secara global yang tercatat di hutan yang Tertebang-dan-Terbakar-17-tahun.
Deskripsi jaringan dan properti struktural
Jaringan menunjukkan jumlah spesies pemakan buah yang berkurang dan ukuran jaringan yang lebih kecil di Hutan yang ditebang dan dibakar selama 17 tahun (Pemakan buah: rata-rata = 28,5 ± SD = 6,5, Ukuran jaringan: rata-rata = 44 ± SD = 6,0) dibandingkan dengan yang tidak terganggu (Pemakan buah: rata-rata = 44,4 ± SD = 8,88, Ukuran jaringan: rata-rata = 59,6 ± SD = 7,12, Tabel 1 , Informasi pendukung). Sebaliknya, tidak terdeteksi adanya perbedaan signifikan pada hutan yang ditebang (Frugivora: rata-rata = 45 ± SD = 4,5, Ukuran jaringan: rata-rata = 63,25 ± SD = 7,25) dan hutan yang ditebang dan dibakar selama 3 tahun (Frugivora: rata-rata = 46,4 ± SD = 4,0, Ukuran jaringan: rata-rata = 67,25 ± SD = 4,62) jika dibandingkan dengan hutan yang tidak terganggu (Tabel 1 ). Demikian pula, jumlah total interaksi juga lebih rendah di hutan yang ditebang dan dibakar selama 17 tahun (rata-rata = 168 ± SD = 65,30), tetapi tidak berbeda secara signifikan antara hutan yang ditebang (rata-rata = 298,75 ± SD = 64,22), dan hutan yang ditebang dan dibakar selama 3 tahun (rata-rata = 239 ± SD = 43,50) jika dibandingkan dengan hutan yang tidak terganggu (rata-rata = 369,40 ± SD = 184,57, Tabel 1 , Informasi pendukung). Selain itu, Hutan yang ditebang dan dibakar selama 17 tahun memiliki jumlah interaksi pemakan buah yang unik yang lebih rendah (rata-rata = 47,25 ± SD = 7,08, Tabel 1 ), sedangkan Hutan yang ditebang (rata-rata = 82,50 ± SD = 11,68), dan Hutan yang ditebang dan dibakar selama 3 tahun (rata-rata = 89,75 ± SD = 15,91) serupa dengan Hutan yang tidak terganggu (rata-rata = 74,00 ± SD = 19,40, Tabel 1 ).
Tabel 1. Hasil dari pengujian model linear umum yang menguji dampak penebangan selektif dan kebakaran hutan, dan upaya pengambilan sampel (memperhitungkan perbedaan intensitas pengambilan sampel di antara kelas hutan dengan bobot kuadrat terkecil, dihitung sebagai akar kuadrat dari jumlah interaksi dibagi dengan akar kuadrat dari ukuran jaringan) pada sifat jaringan pemakan buah hutan Amazon. Kelas hutan dari kiri ke kanan sesuai dengan: Hutan tidak terganggu, Hutan yang ditebang, Hutan yang ditebang dan dibakar selama 17 tahun, dan Hutan yang ditebang dan dibakar selama 3 tahun. Hasil disajikan untuk jumlah spesies tumbuhan, ukuran jaringan spesies pemakan buah (jumlah spesies tumbuhan dan pemakan buah), jumlah interaksi berpasangan yang unik, dan frekuensi interaksi. Interaksi pemakan buah di Hutan tidak terganggu berfungsi sebagai dasar untuk perbandingan dengan semua model lainnya. w AICc – bobot yang memberikan perkiraan probabilitas bahwa model tersebut merupakan pilihan terbaik berdasarkan kriteria AICc, dan interval kepercayaan (CI) yang tidak tumpang tindih dengan nol menunjukkan hasil yang signifikan, disorot dengan huruf tebal
| Metrik jaringan | Model terbaik | dengan AICc | Bahasa Inggris | Bahasa Inggris | 95% CI | dari |
|---|---|---|---|---|---|---|
| Jumlah spesies tanaman | Contoh | 0.73 | ||||
| Mencegat | 3.61 | 0.33 | 2,95, 4,26 | 10.82 | ||
| Contoh | -0,36 | 0.16 | – 0,68, – 0,05 | -2,28 | ||
| Jumlah spesies pemakan buah | Pengolahan Hutan + Pengambilan Sampel | 0.86 | ||||
| Mencegat | 3.12 | 0.30 | 2.53, 3.69 | 10.56 | ||
| Tidak terganggu – Tercatat | 0,08 | 0.10 | -0,12, 0,28 | 0,75 | ||
| Tak Terganggu – Ditebang dan Terbakar 17 Tahun | -0,31 | 0.13 | – 0,56, – 0,05 | -2,36 | ||
| Tidak terganggu – Ditebang dan dibakar selama 3 tahun | 0.20 | 0.12 | -0,03, 0,43 | 1.65 | ||
| Contoh | 0.28 | 0.12 | 0,04, 0,50 | 2.36 | ||
| Ukuran jaringan | Perlakuan | 0.62 | ||||
| Mencegat | Tanggal 4.09 | 0,06 | 3.97, 4.19 | 70.56 | ||
| Tidak terganggu – Tercatat | 0,06 | 0,09 | -0,10, 0,22 | 0,70 | ||
| Tak Terganggu – Ditebang dan Terbakar 17 Tahun | -0,30 | 0.10 | – 0,49, – 0,11 | -3,19 | ||
| Tidak terganggu – Ditebang dan dibakar selama 3 tahun | 0.12 | 0,08 | -0,04, 0,28 | 1.44 | ||
| Pengolahan Hutan + Pengambilan Sampel | 0.38 | |||||
| Mencegat | 3.85 | 0.24 | 3.36, 4.32 | 15.74 | ||
| Tidak terganggu – Tercatat | 0,08 | 0,09 | -0,09, 0,25 | 0.92 | ||
| Tak Terganggu – Ditebang dan Terbakar 17 Tahun | -0,26 | 0.11 | – 0,46, – 0,04 | -2,42 | ||
| Tidak terganggu – Ditebang dan dibakar selama 3 tahun | 0.17 | 0.10 | -0,02, 0,37 | 1.73 | ||
| Contoh | 0.10 | 0.10 | -0,09, 0,29 | 0,99 | ||
| Jumlah interaksi unik | Pengolahan Hutan + Pengambilan Sampel | 0,77 | ||||
| Mencegat | 3.67 | 0.23 | 2.93, 4.25 | 16.29 | ||
| Tidak terganggu – Tercatat | 0.17 | 0,08 | -0,04, 0,42 | 1.15 | ||
| Tak Terganggu – Ditebang dan Terbakar 17 Tahun | -0,32 | 0.10 | – 0,57, – 0,02 | -3,19 | ||
| Tidak terganggu − Ditebang dan dibakar selama 3 tahun | 0.34 | 0,09 | -0,08, 0,63 | 1.72 | ||
| Contoh | 0.26 | 0,09 | 0,02, 0,54 | 2.91 | ||
| Jumlah total interaksi | Pengolahan Hutan + Pengambilan Sampel | 0.82 | ||||
| mencegat | -302,07 | 88.19 | 2.94, 4.00 | -3,43 | ||
| Tidak terganggu − Tercatat | -9,71 | 32.91 | -0,09, 0,29 | -0,30 | ||
| Tak Terganggu − Ditebang dan Terbakar 17 Tahun | -70,13 | 36.10 | – 0,47, – 0,04 | -1,94 | ||
| Tidak terganggu − Ditebang dan dibakar selama 3 tahun | tanggal 16.12 | 37.03 | -0,03, 0,41 | 0.44 | ||
| Contoh | 279.49 | 35.62 | 0,75, 1,17 | 7.85 |
Secara keseluruhan, jaringan pemakan buah secara signifikan lebih modular dan kurang bersarang daripada yang diharapkan oleh model nol sementara signifikansi konektivitas dan spesialisasi bervariasi di antara jaringan (Gbr. 1 , Informasi pendukung). Meskipun demikian, tidak ada efek signifikan dari perawatan dalam metrik jaringan mana pun (Informasi pendukung). 17 jaringan modular terdiri dari 159 modul, berkisar antara 4 hingga 15 di seluruh jaringan. Hutan yang tidak terganggu memiliki jumlah modul terendah (rata-rata = 8,00 ± SD = 3,4), sementara hutan yang ditebang dan dibakar selama 3 tahun memiliki yang tertinggi (rata-rata = 11,50 ± SD = 2,6, Informasi pendukung). Modul individu bervariasi dalam jumlah spesies, berkisar antara dua hingga 45 spesies per modul (rata-rata = 12 ± 6 SD). Jumlah spesies tanaman per modul berkisar dari satu hingga delapan (rata-rata = 2 ± 1 SD), sedangkan spesies pemakan buah berkisar dari satu hingga 33 (rata-rata = 4 ± 1 SD). Di antara semua modul, 39% (n = 48) merupakan modul kecil, yang terdiri dari satu spesies tanaman yang berinteraksi dengan satu pemakan buah.

β-keanekaragaman spesies dan interaksi
Kami menemukan perbedaan tinggi (β sor ) antara area studi untuk spesies tumbuhan (β Plants = 0,94), spesies frugivora (β Frug = 0,86), dan interaksi frugivora (β Int = 0,97, Gambar 2 , Informasi pendukung). Meskipun perbedaan tinggi dalam komposisi spesies dan interaksi juga diamati dalam kelas intra-hutan, perbedaan ini lebih kecil dibandingkan dengan kelas antar-hutan (Informasi pendukung). Pola ini konsisten di semua perbandingan berpasangan untuk tumbuhan, frugivora, dan interaksi (Informasi pendukung). Pergantian spesies adalah pendorong utama perubahan dalam komposisi spesies dan interaksi antara area studi. Pergantian β (β sim ) tinggi untuk spesies tumbuhan (β Plants = 0,92) dan spesies frugivora (β Frug = 0,83), serta untuk interaksi (β Int = 0,97). Sebaliknya, β-nestedness (β nes ) rendah untuk spesies tumbuhan (β Plants = 0,01), spesies pemakan buah (β Frug = 0,04), dan untuk interaksi (β Int = <0,01). Pergantian β spesies juga merupakan komponen utama yang mendorong perbedaan dalam komposisi spesies dan interaksi di dalam dan di antara kelas hutan, dengan β-nestedness tetap rendah secara konsisten (Informasi pendukung).
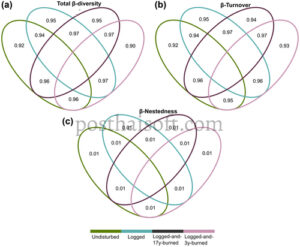
Ketidakmiripan spesies tanaman tinggi, dengan hanya tiga spesies (yaitu Didymopanax morototoni , Laetia procera dan Virola michelii ) yang ditemukan di semua kelas hutan (Informasi pendukung). Untuk frugivora, dari 174 spesies yang tercatat, 68 eksklusif untuk satu kelas hutan sementara 29 spesies (21 burung dan delapan mamalia, atau 16,6% dari semua spesies) hadir di semua kelas hutan (Informasi pendukung). Dari 991 interaksi unik, hanya satu interaksi berpasangan tanaman-frugivora yang tercatat di semua kelas hutan, yaitu Virola michelii dan frugivora terkaitnya, Ramphastos tucanus toucan berleher putih (Informasi pendukung).
Perbedaan spesies dan interaksi tercermin dalam kedua sumbu NMDS dua dimensi ( Gambar 3 ). Daerah studi dalam kelas hutan yang sama menunjukkan distribusi sumbu yang lebih dekat untuk spesies dan interaksi, yang menunjukkan kesamaan yang lebih besar di dalam kelas hutan. Mengenai perbedaan antar kelas hutan, hutan yang ditebang dan dibakar selama 17 tahun menunjukkan skor negatif untuk tumbuhan dan hampir sepenuhnya terpisah dari hutan yang tidak terganggu, yang memiliki skor positif (Gambar 3a ). Untuk spesies pemakan buah, hutan yang ditebang dan dibakar selama 17 tahun berasosiasi positif dengan sumbu NMDS pertama, sedangkan hutan yang tidak terganggu berasosiasi negatif dengannya (Gambar 3b ). Hasil ini menunjukkan komposisi komunitas yang kontras untuk tumbuhan dan pemakan buah. Demikian pula, interaksi pemakan buah di hutan yang ditebang dan dibakar selama 17 tahun menunjukkan skor negatif di sepanjang sumbu pertama, yang kontras dengan hutan yang tidak terganggu, yang berasosiasi dengan skor positif di sepanjang sumbu kedua (Gambar 3c ). Pola-pola ini menunjukkan bahwa hutan yang ditebang dan dibakar selama 17 tahun memiliki perbedaan spesies dan interaksi yang lebih tinggi dibandingkan dengan hutan yang tidak terganggu ( Gbr. 3 ). Sebaliknya, hutan yang ditebang dan ditebang dan dibakar selama 3 tahun menunjukkan hubungan dengan sumbu NMDS yang mirip dengan hutan yang tidak terganggu, yang menunjukkan kesamaan yang lebih besar dalam komposisi spesies tumbuhan dan pemakan buah serta interaksinya ( Gbr. 3 ).

Diskusi
Studi kami menggambarkan interaksi frugivori di seluruh komunitas di hutan Amazon yang tidak terganggu dan yang dimodifikasi oleh manusia. Kami menemukan hilangnya spesies dan interaksi yang signifikan di hutan yang ditebang secara selektif dan dibakar hampir dua dekade lalu, tetapi tidak ada perubahan di hutan yang ditebang secara selektif atau ditebang secara selektif dan dibakar baru-baru ini. Namun, baik penebangan selektif maupun kebakaran hutan tidak mengubah sifat struktural jaringan frugivori, yang tetap sangat modular dan terspesialisasi, tidak bersarang dan terhubung dengan buruk di seluruh kelas hutan. Semua kelas hutan, terlepas dari jenis gangguan antropogenik, menunjukkan tingkat keragaman β spesies dan interaksi yang tinggi. Perbedaan lebih tinggi antara hutan yang tidak terganggu dan yang ditebang secara selektif dan dibakar hampir dua dekade lalu, terutama didorong oleh pergantian spesies dalam komposisi spesies dan interaksi.
Dampak penebangan selektif dan kebakaran terhadap spesies dan interaksinya
Menurut ekspektasi, hutan yang ditebang dan dibakar secara selektif memiliki lebih sedikit spesies dan interaksi dibandingkan dengan hutan yang tidak terganggu. Secara khusus, hutan yang ditebang secara selektif yang terbakar hampir dua dekade lalu menunjukkan jumlah spesies yang lebih rendah, serta jumlah interaksi total dan unik yang lebih sedikit, relatif terhadap hutan yang tidak terganggu. Secara khusus, hutan dengan jenis gangguan seperti itu menunjukkan pengurangan 16% dalam jumlah spesies, penurunan 65,6% dalam jumlah interaksi, dan pengurangan 34,5% dalam interaksi unik, relatif terhadap hutan yang tidak terganggu. Pengurangan spesies dan interaksi ini dapat mengakibatkan hilangnya serangkaian sifat dan sejarah evolusi yang tak tergantikan, yang selanjutnya dapat membahayakan fungsi ekologis yang terkait dengan dinamika hutan (Emer et al. 2019 , Souza et al. 2022 ). Selain itu, tidak adanya spesies kunci, termasuk yang terancam secara global, di hutan yang ditebang dan dibakar secara selektif hampir dua dekade lalu menyoroti ketidakmampuan hutan ini untuk mendukung spesies yang sensitif terhadap gangguan antropogenik. Misalnya, meskipun kami menemukan manakin bermahkota opal dan monyet laba-laba berpipi putih di kelas hutan lain, mereka tidak ada di hutan yang ditebang dan dibakar secara selektif hampir dua dekade lalu. Hasil ini menunjukkan adanya potensi jeda waktu antara penebangan selektif dan kebakaran hutan serta efek berjenjangnya pada spesies dan, selanjutnya, pada interaksi. Periode yang panjang ini kemungkinan mencerminkan waktu respons spesies yang berbeda terhadap gangguan, yang menyoroti bahwa waktu respons sering kali tidak langsung. Misalnya, di Amazon, kematian tanaman akibat kebakaran dapat bertahan hingga delapan tahun setelah kebakaran hutan (Brando et al. 2014 , Silva et al. 2018 ). Sementara tanaman di hutan yang baru saja terbakar mungkin terus menghasilkan buah dan bahkan dapat meningkatkan produksi dalam jangka pendek, kematian pohon yang berkelanjutan kemungkinan menyebabkan penurunan produksi secara keseluruhan (Barlow dan Peres 2006 , Brando et al. 2024 ), yang dapat menjelaskan hilangnya spesies dan interaksi pada akhirnya dalam skala waktu yang lebih panjang (Herrera 1985 , García et al. 2011 ).
Dampak penebangan selektif dan kebakaran pada properti jaringan
Bertentangan dengan hipotesis kami, kami tidak menemukan efek gangguan antropogenik pada struktur jaringan. Semua jaringan pemakan buah sangat modular, yang menunjukkan bahwa interaksi terstruktur menjadi subkelompok yang berbeda. Modularitas dan spesialisasi yang diamati menunjukkan tumpang tindih relung yang rendah dan, akibatnya, redundansi fungsional yang rendah (Cordeiro et al. 2020 , Oliveira et al. 2022 ). Pola ini selanjutnya tercermin oleh variabilitas dalam nilai konektivitas dan tingkat bersarang jaringan yang rendah secara konsisten. Ada tiga alasan potensial dan tidak saling eksklusif untuk kurangnya efek tingkat jaringan yang kuat. Pertama, modularitas dan spesialisasi yang tinggi dapat mencerminkan partisi relung yang kuat di antara spesies dan interaksinya. Dengan menempati relung ekologi yang berbeda, spesies meminimalkan persaingan langsung satu sama lain, yang berpotensi memfasilitasi pembentukan modul yang berbeda dalam jaringan (Ponisio et al. 2019 , Cordeiro et al. 2020 ). Dalam modul-modul ini, spesies berinteraksi lebih sering dengan subset spesies, meningkatkan spesialisasi keseluruhan dalam jaringan. Pemisahan relung seperti itu juga dapat menjelaskan mengapa jaringan mempertahankan struktur modular dan terspesialisasi meskipun komposisi spesies bervariasi di berbagai area studi (Schleuning et al. 2012 , Oliveira et al. 2022 ). Kedua, keanekaragaman tumbuhan dan pemakan buah yang tinggi di hutan Amazon kemungkinan berkontribusi pada pola modular dan terspesialisasi yang muncul yang diamati (Donatti et al. 2011 , Messeder et al. 2020 ). Misalnya, hutan Amazon diketahui menampung spesies tumbuhan yang berbuah beberapa kali per tahun, yang lain yang bereproduksi setiap tahun, dan yang lain lagi yang mungkin memerlukan beberapa tahun di antara episode berbuah (Haugaasen dan Peres 2005 , Bentos et al. 2008 , Mendoza et al. 2018 ). Meskipun demikian, pengambilan sampel kelompok pemakan buah tertentu, termasuk burung dan mamalia, serta perbedaan antara pemakan buah terestrial dan arboreal, juga dapat berkontribusi pada penggabungan struktur modular. Dengan demikian, upaya pengambilan sampel mungkin memiliki dampak kecil pada properti jaringan (Nielsen dan Bascompte 2007 ) – struktur jaringan alternatif dapat muncul dengan upaya pengambilan sampel yang mencakup beberapa tahun. Pendekatan ini dapat menghasilkan representasi yang lebih komprehensif dari semua kemungkinan interaksi (Vázquez et al. 2009 , Vizentin-Bugoni et al. 2016 ). Akhirnya, kurangnya dampak pada properti jaringan dapat mencerminkan keberadaan matriks hutan berkelanjutan yang mengelilingi area studi (Hagen et al. 2012)). Area studi tertanam dalam bentang hutan yang luas dan matriks kaya spesies yang dapat mendukung sifat struktural jaringan, khususnya di hutan yang ditebang secara selektif dan dibakar hanya satu kali. Namun, sementara efeknya mungkin lebih kuat di wilayah dengan tutupan hutan yang lebih sedikit, ini jauh dari pasti karena 1) wilayah studi kami secara luas cocok dengan perubahan sifat tanaman dan penggantian spesies pohon pemenang-pecundang di wilayah hutan yang dimodifikasi manusia lainnya (Pinho et al. 2024 ), 2) efek gangguan hutan cukup kuat untuk memengaruhi komposisi komunitas dan metrik keanekaragaman lainnya untuk tanaman dan pemakan buah, dan interaksi (Barlow dan Peres 2004 , Burivalova et al. 2014 , Carvalho et al. 2022 , Brando et al. 2024 , Grau-Andrés et al. 2024 ).
Dampak penebangan selektif dan kebakaran hutan terhadap keragaman β jaringan
Seperti yang dihipotesiskan, keanekaragaman hayati β tinggi antara area studi dan antara kelas hutan untuk spesies dan interaksi. Namun, alih-alih hutan yang ditebang dan dibakar secara selektif baru-baru ini, hutan yang ditebang dan dibakar secara selektif hampir dua dekade lalu yang menampilkan spesies dan komposisi interaksi yang paling berbeda dibandingkan dengan hutan yang tidak terganggu. Meskipun lebih kecil daripada perbedaan antara kelas hutan, nilai keanekaragaman hayati β yang tinggi juga diamati dalam setiap kelas hutan. Ketahanan keanekaragaman hayati yang tinggi pada famili burung tertentu (misalnya Cotingidae, Pipridae, Ramphastidae dan Thraupidae) di seluruh area hutan dapat berkontribusi pada pemeliharaan nilai keanekaragaman hayati β yang tinggi (Moura et al. 2013 , Acevedo-Quintero et al. 2020 ). Selain itu, keanekaragaman hayati yang cukup besar pada famili tumbuhan tertentu seperti Melostomataceae, yang menghasilkan buah-buahan kecil yang penting bagi banyak spesies pemakan buah, juga dapat memainkan peran penting dalam mempertahankan nilai keanekaragaman hayati β yang tinggi (Messeder et al. 2021 ). Peningkatan keanekaragaman β yang diamati mungkin mencerminkan peningkatan fungsionalitas sistem (Mori et al. 2018 ), menyoroti kompleksitas besar wilayah megadiverse (Valiente-Banuet et al. 2015 , Solar et al. 2015 , Bruno et al. 2024 ).
Nilai β-diversity yang tinggi sebagian besar dikaitkan dengan β-turnover, daripada β-nestedness, yang menunjukkan bahwa penebangan selektif dan kebakaran hutan memainkan peran penting dalam membentuk komposisi komunitas frugivori (Mestre et al. 2013 ). Hal ini kemungkinan merupakan konsekuensi dari perubahan struktur hutan setelah gangguan antropogenik (Mestre et al. 2013 , Morante-Filho et al. 2016 ). Misalnya, tajuk yang lebih terbuka akibat penebangan selektif dan kebakaran hutan dapat meningkatkan keberadaan spesies burung tepi dan non-hutan seperti Megarynchus pitangua boat-billed flycatcher dan Pitangus sulphuratus great kiskadee, yang menyebabkan peningkatan pergantian spesies baik spesies maupun interaksi (Barlow dan Peres 2008 , Lees et al. 2013 ). Selain itu, seiring berjalannya waktu, spesies tanaman pionir mulai merekrut di hutan yang dimodifikasi manusia, mengubah komposisi komunitas dan karenanya berkontribusi pada pergantian β yang tinggi (Barlow dan Peres 2008 , Carvalho et al. 2022 ). Terakhir, waktu yang berlalu sejak peristiwa gangguan juga dapat memainkan peran penting dalam mempertahankan nilai keragaman β, karena dapat mengakibatkan variasi yang lebih besar dalam jalur suksesi (Norden et al. 2015 ).
Penutup
Studi kami memberikan bukti baru bahwa gangguan antropogenik menyederhanakan interaksi pemakan buah di hutan Amazon. Melalui upaya pengambilan sampel yang ekstensif selama 31.484 jam untuk memantau interaksi arboreal dan terestrial, kami menemukan bahwa penebangan selektif dan kebakaran hutan menyebabkan hilangnya spesies dan interaksi yang signifikan, dengan hutan yang dimodifikasi manusia menunjukkan komposisi taksonomi yang berbeda dibandingkan dengan hutan yang tidak terganggu. Temuan ini menggarisbawahi peran penting penebangan selektif dan kebakaran dalam membentuk interaksi ekologis, terutama ketika bertindak secara sinergis. Sebaliknya, struktur jaringan tidak terpengaruh oleh gangguan antropogenik ini, yang menunjukkan kemampuan komunitas interaksi untuk mempertahankan fungsi ekologis bahkan di hutan yang dimodifikasi manusia. Namun, hilangnya dan perubahan interaksi yang berkelanjutan, dikombinasikan dengan peningkatan frekuensi dan tingkat keparahan kebakaran yang diprediksi karena perubahan iklim, serta penebangan selektif yang sedang berlangsung, dapat mengancam ketahanan hutan tropis. Spesies tumbuhan dan pemakan buah saja mungkin tidak cukup untuk mempertahankan fungsionalitas ekosistem, karena sebagian besar spesies tidak beradaptasi dengan kebakaran hutan, peningkatan suhu, heterogenitas iklim, dan perubahan lingkungan terkait, seperti berkurangnya luas hutan. Masih belum jelas bagaimana interaksi pemakan buah menanggapi kebakaran hutan dalam jangka panjang (yaitu selama beberapa dekade) di seluruh Amazon – jika hutan yang terbakar terus kehilangan interaksi pemakan buah atau mencapai keadaan stabil dengan jaringan pemakan buah yang disederhanakan, pemulihan hutan ini akan sangat terdampak.