
ABSTRAK
Kekurangan zat besi (Fe) merupakan kendala kritis pada ketahanan pangan global, khususnya yang mempengaruhi tanaman hortikultura bernilai tinggi seperti stroberi ( Fragaria × ananassa ). Studi ini meneliti peran melatonin dan pensinyalan hidrogen sulfida (H 2 S) dalam mengurangi stres kekurangan Fe dengan meningkatkan bioavailabilitas Fe dan meningkatkan ketahanan tanaman. Tanaman stroberi dibudidayakan dalam kondisi cukup Fe dan kekurangan Fe dan diobati dengan 100 μM melatonin dan 3 mM dl-propargylglycine (PAG), penghambat L-sistein desulfhidrase (L-DES), yang mengatur produksi H 2 S. Kekurangan Fe secara signifikan mengurangi kandungan klorofil dan efisiensi fotosintesis sambil meningkatkan penanda stres oksidatif seperti hidrogen peroksida (H 2 O 2 ), malondialdehid (MDA), dan kebocoran elektrolit (EL). Aplikasi melatonin meringankan efek kekurangan Fe dengan meningkatkan pemanfaatan Fe, merangsang aktivitas L-DES, dan meningkatkan produksi H 2 S. Melatonin juga meningkatkan pertahanan antioksidan dengan meningkatkan aktivitas superoksida dismutase (SOD), katalase (CAT), dan peroksidase (POD), serta mempertahankan dinamika redoks askorbat-glutathione (AsA-GSH). Penambahan 3 mM PAG menghambat aktivitas L-DES, yang mengakibatkan penurunan kadar H 2 S dan manfaat yang diinduksi melatonin, yang menggarisbawahi peran penting sintesis H 2 S yang dimediasi L-DES. Meskipun terdapat PAG, aplikasi bersama natrium hidrosulfida (NaHS) 0,2 mM dan melatonin memulihkan bioavailabilitas Fe, pertumbuhan, dan kapasitas antioksidan, yang menunjukkan interaksi sinergis antara melatonin dan H 2 S. Studi ini menyoroti potensi melatonin dan pensinyalan H 2 S untuk meningkatkan homeostasis Fe dan mengurangi stres oksidatif pada tanaman yang kekurangan Fe. Temuan ini menawarkan strategi untuk meningkatkan ketahanan dan produktivitas tanaman di tanah yang kekurangan nutrisi, sehingga mendorong pertanian berkelanjutan dan ketahanan pangan global.
1 Pendahuluan
Besi (Fe) adalah mikronutrien esensial bagi tanaman, memainkan peran kunci dalam sintesis klorofil, reaksi enzimatik, dan respirasi. Meskipun Fe melimpah di kerak bumi, bioavailabilitasnya bagi tanaman sering dibatasi oleh sifat tanah yang mengurangi kelarutan dan mobilitasnya (Colombo et al. 2014 ). Bioavailabilitas Fe sangat dipengaruhi oleh faktor-faktor seperti potensial redoks tanah, kadar pH, bahan organik, dan komposisi mineral lempung. Di tanah alkali dan berkapur, Fe umumnya membentuk hidroksida dan oksihidroksida yang tidak larut, yang secara signifikan membatasi penyerapannya oleh tanaman (Riaz et al. 2020 ). Kekurangan Fe sangat parah di tanah yang teroksigenasi, di mana Fe 3+ , bentuk yang dominan, memiliki kelarutan yang jauh lebih rendah daripada Fe 2+ , yang lebih mudah diserap oleh tanaman.
Konsekuensi dari defisiensi Fe sangat besar, yang menyebabkan klorosis, pertumbuhan terhambat, dan penurunan produktivitas pertanian, yang berdampak buruk pada hasil dan kualitas tanaman (Duralija et al. 2021 ; Roychoudhury et al. 2025 ). Di luar pertanian, defisiensi Fe memiliki implikasi yang signifikan terhadap nutrisi manusia, khususnya di wilayah-wilayah yang didominasi oleh pola makan nabati, yang berkontribusi terhadap meluasnya anemia defisiensi Fe dan risiko kesehatan terkait (Mantadakis et al. 2020 ). Oleh karena itu, mengatasi defisiensi Fe merupakan prioritas global untuk meningkatkan keberlanjutan pertanian dan kesehatan manusia (Mahawar et al. 2023 ).
Tanaman stroberi ( Fragaria × ananassa ) sangat sensitif terhadap defisiensi Fe, sehingga menjadikannya model ideal untuk mempelajari bioavailabilitas Fe dan respons stres. Pada stroberi, defisiensi Fe biasanya menyebabkan klorosis interveinal, berkurangnya biomassa, dan kualitas buah yang menurun, terutama di tanah berkapur dengan kelarutan Fe yang rendah (Duralija et al. 2021 ). Meningkatkan kandungan Fe dan ketahanan tanaman stroberi tidak hanya meningkatkan produktivitas tanaman tetapi juga berpotensi meningkatkan nutrisi manusia, mengingat popularitas global buah ini dan pentingnya dalam makanan (Sabahat et al. 2021 ).
Metode tradisional untuk mengatasi defisiensi Fe meliputi amandemen tanah, agen khelasi, dan pemuliaan varietas tanaman yang efisien dalam Fe (Colombo et al. 2014 ; Ali et al. 2023 ). Selain itu, memanfaatkan mikroba bermanfaat yang menghasilkan siderofor untuk meningkatkan kelarutan Fe telah menunjukkan hasil yang menjanjikan (Zhang et al. 2023 ). Namun, pendekatan ini sering kali memiliki keterbatasan dalam hal biaya, keberlanjutan lingkungan, dan skalabilitas. Baru-baru ini, fitohormon seperti melatonin telah menarik perhatian karena kemampuannya untuk memodulasi pertumbuhan tanaman dan respons stres di bawah tekanan abiotik, termasuk defisiensi nutrisi (Nawaz et al. 2020 ; Zhang, Wang, et al. 2022 ). Studi telah menunjukkan bahwa melatonin meningkatkan penyerapan Fe, mengatur homeostasis Fe, dan mengurangi stres oksidatif yang terkait dengan defisiensi Fe, menghadirkan pendekatan inovatif untuk mengatasi tantangan ini (Zhou et al. 2016 ; Ahammed et al. 2020 ).
Bidang penelitian lain yang menjanjikan melibatkan H 2 S, molekul pensinyalan yang disintesis dalam tanaman melalui aktivitas enzimatik L-DES. H 2 S yang pernah dianggap sebagai gas beracun, sekarang diakui karena fungsi pentingnya dalam mengendalikan perkecambahan biji, pertumbuhan akar, dan respons stres abiotik (Shen et al. 2019 ; Zhang et al. 2021 ). Penelitian terbaru telah menunjukkan bahwa H 2 S meningkatkan kelarutan dan penyerapan Fe dengan mengatur proses khelasi Fe dan meningkatkan fungsionalitas akar dalam kondisi kekurangan Fe (Chen et al. 2015 , 2020 ). Lebih jauh, penelitian mengusulkan interaksi integratif antara melatonin dan H 2 S, di mana melatonin memodulasi aktivitas L-DES, dengan demikian memengaruhi produksi H 2 S dan memperkuat efek perlindungannya pada tanaman yang menghadapi stres lingkungan (Li et al. 2024 ; Ghorbani et al. 2024 ).
Meskipun peran H 2 S dalam fisiologi tanaman telah ditetapkan, jalur pasti yang dilalui melatonin dalam memengaruhi aktivitas L-DES dan selanjutnya memodulasi produksi H 2 S dalam kondisi kekurangan Fe masih belum dieksplorasi secara memadai. Memahami bagaimana perubahan yang dimediasi melatonin dalam aktivitas L-DES memengaruhi kadar H 2 S, dan akibatnya penyerapan dan penggunaan Fe pada tanaman, sangat penting untuk merumuskan pendekatan yang ditargetkan guna meningkatkan bioavailabilitas Fe. Penelitian ini bertujuan untuk menyelidiki pengaruh melatonin pada aktivitas L-DES untuk memodulasi produksi H 2 S sebagai reaksi terhadap kekurangan zat besi dan untuk mengevaluasi bagaimana perubahan kadar H 2 S memengaruhi kelarutan, penyerapan, dan distribusi Fe pada tanaman stroberi. Kami berhipotesis bahwa melatonin meningkatkan bioavailabilitas Fe pada tanaman stroberi yang kekurangan Fe dengan meningkatkan aktivitas L-DES, sehingga meningkatkan produksi H 2 S. Hal ini, pada gilirannya, meningkatkan kelarutan, penyerapan, dan distribusi Fe, sehingga mengurangi efek merugikan dari kekurangan Fe.
2 Bahan dan Metode
2.1 Budidaya dan Perawatan Tanaman Stroberi
Percobaan rumah kaca dalam kondisi terkendali dilakukan menggunakan kultivar stroberi ‘Camarosa’ ( Fragaria × ananassa Duch.). Tanaman dibudidayakan dalam kondisi terkendali, dengan siklus suhu 13 jam pada malam hari (15°C) dan 11 jam pada siang hari (25°C), dan kelembaban relatif 65%–70%. Tanaman stroberi dengan diameter tajuk 0,8–1,0 cm ditanam dalam wadah 2 L yang diisi perlit. Wadah memiliki lubang drainase dan ditutup dengan lembaran plastik hitam untuk mencegah pertumbuhan alga dan mengurangi penguapan.
Tanaman diairi setiap 2 hari dengan 0,1–1,0 L larutan nutrisi Hoagland (Kaya dan Ashraf 2015 ), disesuaikan menurut tahap pertumbuhan untuk mempertahankan hidrasi yang memadai. pH larutan dipertahankan pada 5,5 menggunakan penyangga berbasis fosfat. Pencahayaan buatan digunakan untuk mempertahankan periode cahaya 11 jam selama percobaan.
Tujuh hari setelah penanaman, setengah dari tanaman diperlakukan dengan 0,1 mM melatonin dan 0,01% Tween-20, yang digunakan sebagai surfaktan untuk meningkatkan daya rekat daun. Perlakuan ini diterapkan setiap dua hari selama 6 hari. Kelompok tanaman kontrol diperlakukan dengan semprotan yang mengandung 0,01% Tween-20 dalam air deionisasi suling. Sebelum induksi stres, bibit terpilih diinkubasi selama 12 jam dalam 0,3 mM dL-propargylglycine (PAG), penghambat L-DES, untuk menekan sintesis H 2 S endogen. Selama periode stres, tanaman yang menerima perlakuan melatonin dan PAG juga diberikan 0,2 mM NaHS melalui larutan nutrisi untuk memeriksa peran H 2 S dalam mengurangi stres. Gambar 1A menguraikan jadwal dan perlakuan eksperimen.

Setiap perlakuan dalam rancangan percobaan diulang tiga kali, dengan lima tanaman per ulangan. Stres defisiensi Fe disebabkan oleh drainase terkontrol dari larutan nutrisi berlebih untuk mencegah akumulasi Fe di zona akar. Setelah 14 hari paparan stres, enam tanaman per perlakuan (dua tanaman per ulangan) dikumpulkan untuk analisis zat besi daun dan akar, sedangkan tiga tanaman sisanya per ulangan (total sembilan tanaman per perlakuan) digunakan untuk analisis fisiologis dan biokimia.
2.2 Kandungan Klorofil dan Fluoresensi Klorofil
Penilaian kandungan klorofil dilakukan dengan mengikuti metode Strain et al. ( 1971 ). Bahan daun segar (1,0 g) dihomogenkan dalam 5 mL aseton 90% ( v / v ), dan ekstrak disaring dan disimpan dalam tabung yang terlindungi cahaya. Absorbansi pada 663,5 nm dan 645 nm diukur menggunakan spektrofotometer untuk menghitung konsentrasi klorofil a dan b.
Fluoresensi klorofil dinilai dengan menentukan rasio Fv/Fm menggunakan fluorometer klorofil Mini-PAM (Walz, Jerman). Daun disimpan dalam gelap selama 30 menit untuk beradaptasi sebelum pengukuran, dengan enam pembacaan per replikasi dirata-ratakan untuk setiap perlakuan.
2.3 Penilaian Kandungan Besi Total dan Aktif, Bersama dengan Aktivitas Ferric-Chelate Reductase (FCR)
Konsentrasi total Fe dalam jaringan daun diukur menurut metode yang dijelaskan oleh Lei et al. ( 2014 ). Setelah dicuci dalam 1,5 mM CaCl2 , daun dicerna dengan larutan HNO3 dan HClO4 4:1 (v/v) , dan dianalisis menggunakan spektrometer Inductively Coupled Plasma (ICP) (PerkinElmer Optima 5300 DV).
Kandungan zat besi aktif dalam jaringan daun ditentukan dengan mengikuti prosedur yang dijelaskan oleh Gao dan Shi ( 2007 ). Daun segar (2,0 g) dicincang halus, dihomogenkan dalam 20 mL HCl 1 M, dan dikocok selama 5 jam, menurut metode Takker dan Kaur ( 1984 ). Setelah penyaringan, konsentrasi zat besi aktif diukur menggunakan spektrometer ICP yang sama.
Aktivitas reduktase ferri-kelat akar dievaluasi menurut Xu et al. ( 1998 ). Akar dibilas dengan larutan jenuh CaSO4 selama 5 menit dan dibersihkan dengan air deionisasi sebelum dipindahkan ke larutan yang terdiri dari 0,1 mM ferri-EDTA dan 0,4 mM 2,2′-bipiridil, yang diatur pada pH 5,0. Setelah 2 jam dalam kondisi eksperimen, aktivitas FCR dinilai dengan mengukur absorbansi senyawa Fe(II)-bipiridil pada 562 nm.
2.4 Pengukuran Aktivitas L-DES dan Produksi H 2 S
Aktivitas L-DES ditentukan menggunakan metode Riemenschneider et al. ( 2005 ) dengan modifikasi. 5 g sampel daun dihomogenkan dalam nitrogen cair dan diekstraksi dengan buffer Tris-HCl 20 mM (pH 8,0). Aktivitas enzim diuji dengan mengukur absorbansi pada 667 nm setelah reaksi dengan L-sistein, FeCl3 , dan N,N-dimetil-p-fenilendiamin.
Sampel jaringan daun sebanyak 0,3 g digerus dalam buffer fosfat (pH 7,0) yang mengandung EDTA 10 mM. Supernatan direaksikan dengan 5,5′-dithiobis (asam 2-nitrobenzoat), dan absorbansi pada 412 nm direkam untuk menentukan konsentrasi H 2 S (Nashef et al. 1977 ).
2.5 Pengukuran Penanda Stres Oksidatif
Hidrogen peroksida diukur menggunakan metode Velikova et al. ( 2000 ). Daun segar dihomogenkan dalam 1% TCA, dan absorbansi pada 390 nm diukur setelah reaksi dengan kalium iodida.
Kandungan malondialdehid, penanda peroksidasi lipid, diukur menggunakan metode yang dijelaskan oleh Weisany et al. ( 2012 ). Ekstrak daun direaksikan dengan asam tiobarbiturat (TBA) dan absorbansi pada 532 nm dicatat. Kebocoran elektrolit (EL) dievaluasi seperti yang dijelaskan oleh Dionisio-Sese dan Tobita ( 1998 ) dengan mengukur konduktivitas listrik sebelum dan setelah cakram daun diautoklaf.
2.6 Aktivitas Enzim Antioksidan dan Kandungan Protein Larut
Homogenisasi sampel daun segar (0,5 g) dilakukan dalam buffer natrium fosfat 50 mM yang mengandung 1% polivinilpirolidon (PVP). Setelah disentrifugasi homogenat pada 20.000 × g selama 15 menit pada suhu 4°C, supernatan yang dihasilkan digunakan untuk mengukur aktivitas CAT, SOD, dan POD.
Penurunan absorbansi pada 240 nm dipantau untuk mengukur aktivitas katalase, yang menunjukkan kerusakan hidrogen peroksida, mengikuti metode Chance dan Maehly ( 1955 ). Aktivitas peroksidase ditentukan dengan melacak peningkatan absorbansi pada 470 nm karena oksidasi guaiacol dengan adanya hidrogen peroksida. Mengikuti metode Van Rossum et al. ( 1997 ), aktivitas superoksida dismutase dinilai berdasarkan efeknya pada penghambatan reduksi NBT fotokimia. Aktivitas enzim dihitung berdasarkan jumlah yang diperlukan untuk penghambatan 50% reduksi NBT.
Untuk penentuan protein, digunakan uji Bradford (Bradford 1976 ). Buffer fosfat digunakan untuk menghomogenkan jaringan daun segar, diikuti dengan sentrifugasi. Reagen Coomassie Brilliant Blue ditambahkan ke supernatan yang diencerkan, dan absorbansi dicatat untuk mengukur kandungan protein.
2.7 Kuantifikasi Askorbat (AsA) dan Glutathione (GSH)
Untuk mengukur kadar AsA dan GSH, 500 mg jaringan daun segar dicampur menjadi campuran homogen dalam 3 mL larutan dingin yang mengandung 5% asam metafosfat dan EDTA. Sentrifugasi homogenat dilakukan pada 11.500 × g selama 12 menit pada suhu 4°C, dan supernatan dipisahkan dan disimpan untuk analisis selanjutnya. Kandungan AsA ditentukan dengan menetralkan 0,4 mL alikuot ekstrak dengan buffer dan mengukur absorbansi pada 265 nm dengan adanya askorbat oksidase (Huang et al. 2005 ). Kadar GSH dan GSSG diukur mengikuti protokol yang melibatkan penetralan 0,4 mL alikuot ekstrak dengan buffer. GSH diukur dengan mengamati perubahan absorbansi yang sesuai dengan reaksi spesifik (Yu et al. 2003 ). Untuk GSSG, GSH dalam sampel diderivatisasi untuk memungkinkan kuantifikasi selektif.
2.8 Persiapan Ekstrak Tumbuhan Mentah dan Uji Enzim
1,0 mL buffer fosfat (pH 7,0) yang mengandung KCl, AsA, β-merkaptoetanol, dan gliserol digunakan untuk menghomogenkan 0,5 g sampel daun selama 10 menit dalam lumpang dan alu. Supernatan dikumpulkan untuk uji aktivitas enzim berikutnya setelah sentrifugasi homogenat pada 10.000 × g .
Mengikuti prosedur yang diuraikan oleh Nakano dan Asada ( 1981 ), aktivitas askorbat peroksidase (APX) dinilai. Campuran reaksi yang mencakup buffer fosfat (pH 7,0), AsA, dan EDTA dilengkapi dengan ekstrak enzim 0,7 mL. Penambahan H 2 O 2 memicu dimulainya reaksi, dan penurunan absorbansi pada 290 nm diamati selama 1 menit.
Sesuai dengan Hossain et al. ( 2010 ), aktivitas monodehidroaskorbat reduktase (MDHAR) dinilai. Larutan uji terdiri dari ekstrak enzim, askorbat oksidase, NADPH, buffer Tris (pH 7,5), dan AsA. Reaksi dimulai dengan penambahan H 2 O 2 , dan absorbansi diukur pada 340 nm selama 1 menit.
Aktivitas dehidroaskorbat reduktase (DHAR) dinilai menurut Nakano dan Asada ( 1981 ). Absorbansi diukur pada 265 nm setelah larutan enzim digabungkan dengan larutan reaksi yang mengandung GSH, DHA, dan buffer fosfat (pH 7,0).
Metode Hossain dkk. ( 2010 ) digunakan untuk mengevaluasi aktivitas glutathione reduktase (GR). Perubahan absorbansi pada 340 nm diukur setelah ekstrak enzim dimasukkan ke dalam campuran reaksi yang mengandung buffer fosfat (pH 7,0), EDTA, GSSG, dan NADPH.
2.9 Kadar Metilglikosal (MG) dan Uji Sistem Glioksalase
Kadar metilglioksal (MG) ditentukan dengan mengekstraksi 500 mg jaringan daun dalam asam perklorat 5%, diikuti dengan sentrifugasi pada 11.000 × g selama 10 menit pada suhu 4°C. Supernatan yang dihasilkan diperlakukan dengan arang untuk dekolorisasi dan dinetralkan dengan kalium karbonat jenuh pada suhu 25°C. Untuk menyiapkan campuran reaksi, ekstrak diencerkan hingga 1 mL dengan larutan N-asetil-l-sistein dan natrium dihidrogen fosfat dalam rasio 24:25:1. Setelah 10 menit, produk akhir, N-asetil-S-(1-hidroksi-2-okso-propil) sistein, diukur pada 288 nm menggunakan spektrofotometer (Wild et al. 2012 ).
Aktivitas glioksalase I (Gly I) dinilai menurut Hasanuzzaman et al. ( 2014 ). Uji enzim dilakukan dalam larutan yang mengandung buffer kalium fosfat (pH 7,0), GSH, magnesium sulfat, dan MG. Absorbansi dipantau pada 240 nm untuk mengukur aktivitas enzim.
Aktivitas glioksalase II (Gly II) diukur dengan cara yang sama menggunakan metode referensi yang sama. Campuran reaksi meliputi Sd-laktoilglutathione (SLG), DTNB, dan penyangga Tris (pH 7,2), dengan absorbansi yang direkam pada 240 nm untuk mengukur aktivitas Gly II.
2.10 Analisis Statistik
Evaluasi statistik pengaturan parameter dilakukan dengan ANOVA dua arah. Hasil disajikan sebagai nilai rata-rata beserta standar error yang sesuai. Desain eksperimen menggunakan struktur bersarang, dengan kadar besi sebagai faktor plot utama dan perlakuan kimia sebagai subplot. Ambang signifikansi 5% ditetapkan untuk ANOVA, dan analisis post hoc dilakukan menggunakan uji Tukey untuk mengidentifikasi perbedaan signifikan antar kelompok. Batang dengan huruf yang identik pada gambar menunjukkan tidak ada perbedaan signifikan antar kelompok, sedangkan batang dengan huruf yang berbeda menunjukkan perbedaan signifikan secara statistik.
3 Hasil
3.1 Efek Melatonin dan NaHS pada Stroberi Gejala yang Terlihat pada Kekurangan Zat Besi
Tanaman stroberi yang mengalami defisiensi Fe menunjukkan klorosis interveinal pada daun muda dan pertumbuhan terhambat, yang menunjukkan gangguan penyerapan Fe dan bioavailabilitas. Aplikasi melatonin meringankan gejala-gejala ini dengan mengurangi keparahan klorosis dan meningkatkan pertumbuhan tanaman secara keseluruhan (Gambar 1B ). Namun, ketika PAG, penghambat L-DES, diperkenalkan, efek menguntungkan melatonin berkurang drastis. Temuan ini menunjukkan bahwa sintesis H 2 S yang dimediasi L-DES sangat penting bagi peran melatonin dalam mengurangi defisiensi Fe dan meningkatkan kesehatan tanaman.
Menariknya, penggunaan NaHS, donor H 2 S eksogen, bersama melatonin dan PAG mengembalikan efek pemacu pertumbuhan dan meringankan gejala klorosis. Temuan ini menggarisbawahi peran jalur pensinyalan H 2 S dalam memediasi interaksi antara melatonin dan aktivitas L-DES. NaHS tampaknya meningkatkan kadar H 2 S secara independen dari L-DES, mengompensasi penghambatan yang disebabkan oleh PAG dan memulihkan homeostasis Fe.
3.2 Dampak Melatonin dan NaHS terhadap Kandungan Klorofil dan Efisiensi Fotosintesis pada Kekurangan Zat Besi
Kekurangan zat besi secara signifikan mengganggu kinerja fotosintesis pada tanaman stroberi, yang menyebabkan penurunan klorofil a (Chl a), klorofil b (Chl b), dan efisiensi kuantum maksimum fotosistem II (Fv/Fm). Dibandingkan dengan tanaman yang cukup Fe, Chl a, Chl b, dan Fv/Fm menurun masing-masing sebesar 45%, 21%, dan 20% ( p ≤ 0,05) (Gambar 2A–C ). Penurunan ini mencerminkan gangguan sintesis klorofil dan efisiensi fotokimia, yang merupakan gejala khas stres kekurangan Fe.
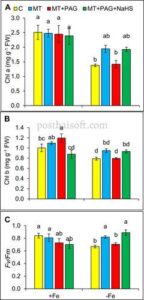
Ketika melatonin diaplikasikan dalam kombinasi dengan PAG, penghambat L-DES, efek perlindungan melatonin berkurang secara signifikan. Kadar Chl a dan Chl b masing-masing berkurang sebesar 27% dan 16%, sementara Fv/Fm hanya menunjukkan peningkatan sebesar 14%, dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati ( p ≤ 0,05). Penurunan ini menyoroti peran penting sintesis H 2 S yang dimediasi L-DES dalam kemampuan melatonin untuk meningkatkan kandungan klorofil dan efisiensi fotosintesis.
Menariknya, penggunaan NaHS, donor H 2 S eksogen, bersama melatonin dan PAG, memulihkan efek menguntungkan yang sebelumnya diamati dengan pengobatan melatonin saja. Dalam pengobatan melatonin + PAG + NaHS, Chl an meningkat sebesar 40%, Chl b sebesar 18%, dan Fv/Fm sebesar 33%, dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati ( p ≤ 0,05). Peningkatan ini menunjukkan bahwa NaHS secara efektif mengompensasi penghambatan sintesis H 2 S endogen yang disebabkan oleh PAG, sehingga memungkinkan melatonin untuk mempertahankan peran protektifnya dalam kondisi kekurangan Fe.
3.3 Melatonin Meningkatkan Pemanfaatan Zat Besi dan Respon Stres melalui Aktivitas L-DES dan Modulasi Sinyal Hidrogen Sulfida
Melatonin memainkan peran kunci dalam mengatur metabolisme Fe dan respons stres pada tanaman stroberi yang kekurangan Fe (Gambar 3A–E ). Melatonin secara signifikan meningkatkan efisiensi penggunaan Fe dengan meningkatkan kadar Fe yang tersedia secara hayati (aktif) tanpa mengubah total kandungan Fe di akar atau daun. Secara khusus, perlakuan melatonin meningkatkan Fe aktif di daun sebesar 69,5% dan di akar sebesar 24,5%, dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati. Peningkatan Fe aktif ini menyoroti peran melatonin dalam memfasilitasi kelarutan Fe dan distribusinya dalam jaringan tanaman.
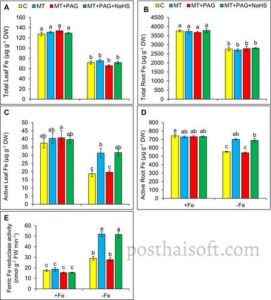
Mekanisme utama yang mendasari efek ini adalah stimulasi aktivitas ferric-chelate reductase (Fe III -reductase) yang diinduksi melatonin, yang memainkan peran penting dalam mereduksi zat besi ferric (Fe 3+ ) menjadi bentuk ferrous yang lebih mudah larut (Fe 2+ ), sehingga meningkatkan penyerapan dan homeostasis Fe. Pada tanaman yang diobati dengan melatonin, aktivitas Fe III -reductase meningkat sebesar 103,8%, dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati, yang menggarisbawahi kemampuan melatonin untuk meningkatkan proses enzimatik yang diperlukan untuk perolehan Fe dalam kondisi stres nutrisi.
Namun, efek menguntungkan melatonin berkurang secara signifikan dengan adanya PAG, penghambat L-DES yang menekan produksi H 2 S endogen . Dalam perlakuan melatonin + PAG, kadar Fe aktif di daun dan akar, serta aktivitas Fe III -reduktase, berkurang, dibandingkan dengan tanaman yang diberi perlakuan melatonin. Penurunan ini menekankan peran penting sintesis H 2 S yang dimediasi L-DES dalam memfasilitasi kemampuan melatonin untuk meningkatkan perolehan dan pemanfaatan Fe.
Ko-aplikasi NaHS dengan melatonin dan PAG memulihkan kadar Fe aktif dan aktivitas Fe III -reduktase. Kadar Fe aktif pada daun dan akar, serta aktivitas Fe III -reduktase, dipulihkan ke kadar yang sebanding dengan yang diamati pada tanaman yang diberi melatonin. Hal ini menunjukkan bahwa suplementasi H 2 S eksogen dapat mengimbangi penghambatan sintesis H 2 S endogen, yang memungkinkan melatonin mempertahankan efek positifnya pada metabolisme Fe dan pengurangan stres.
3.4 Melatonin dan Hidrogen Sulfida Mengurangi Stres Oksidatif yang Disebabkan oleh Kekurangan Zat Besi pada Tanaman Stroberi
Kekurangan zat besi pada tanaman stroberi menyebabkan peningkatan stres oksidatif yang signifikan, sebagaimana dibuktikan oleh peningkatan kadar H 2 O 2 , MDA, dan kebocoran elektrolit (EL). Dibandingkan dengan tanaman yang cukup Fe, kadar H 2 O 2 , kandungan MDA, dan EL meningkat masing-masing sebesar 268%, 248%, dan 141% (Gambar 4A–C ). Temuan ini menunjukkan akumulasi spesies oksigen reaktif (ROS) yang substansial, peroksidasi lipid, dan hilangnya integritas membran, yang semuanya merupakan indikator utama stres oksidatif yang disebabkan oleh kekurangan Fe.
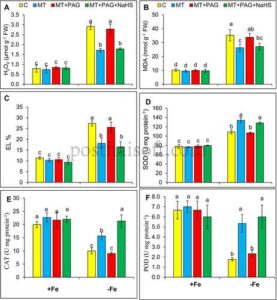
Penambahan PAG, penghambat L-DES, secara signifikan mengurangi efek perlindungan melatonin. Dibandingkan dengan tanaman yang diobati dengan melatonin, aplikasi PAG menghasilkan peningkatan H 2 O 2 sebesar 62% , peningkatan MDA sebesar 27%, dan peningkatan EL sebesar 39% dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati. Penurunan efikasi melatonin ini menyoroti peran penting produksi H 2 S yang dimediasi L-DES dalam fungsi antioksidan melatonin. Penghambatan L-DES mengganggu sintesis H 2 S endogen, yang menyebabkan gangguan detoksifikasi ROS dan eksaserbasi stres oksidatif dalam kondisi kekurangan Fe.
Menariknya, penggunaan NaHS, donor H 2 S eksogen, bersama melatonin dan PAG memulihkan sebagian efek perlindungan melatonin. Kadar H 2 O 2 menurun hingga 38%, kadar MDA hingga 19%, dan EL hingga 36% dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati. Temuan ini menunjukkan bahwa NaHS mengisi kembali kadar H 2 S, yang menangkal efek penghambatan PAG pada aktivitas L-DES. Hasilnya, penambahan NaHS mengembalikan kemampuan melatonin untuk mengurangi akumulasi ROS, peroksidasi lipid, dan kerusakan membran sel.
3.5 Melatonin dan Hidrogen Sulfida Meningkatkan Pertahanan Antioksidan pada Tanaman yang Kekurangan Zat Besi
Kekurangan zat besi mengganggu sistem pertahanan antioksidan pada tanaman, yang menyebabkan perubahan signifikan dalam aktivitas enzim utama seperti SOD, CAT, dan POD (Gambar 4D–F ). Secara khusus, kekurangan Fe menyebabkan peningkatan aktivitas SOD sebesar 40%, yang menunjukkan respons kompensasi terhadap stres oksidatif. Namun, aktivitas CAT dan POD menurun secara signifikan masing-masing sebesar 50% dan 73%, yang mencerminkan berkurangnya kapasitas untuk mendetoksifikasi hidrogen peroksida dan mengelola stres oksidatif dalam kondisi kekurangan Fe. Penurunan signifikan dalam aktivitas CAT dan POD lebih lanjut menunjukkan kapasitas antioksidan yang sangat terganggu dalam kekurangan Fe.
Aplikasi melatonin dalam kondisi kekurangan Fe secara signifikan meningkatkan aktivitas enzim antioksidan ini, sehingga meningkatkan manajemen stres oksidatif. Perlakuan melatonin menyebabkan peningkatan 23% dalam aktivitas SOD dan peningkatan substansial dalam aktivitas CAT dan POD masing-masing sebesar 56% dan 201%, dibandingkan dengan tanaman kekurangan Fe yang tidak diobati. Temuan ini menunjukkan bahwa melatonin meningkatkan aktivitas enzim antioksidan, khususnya CAT dan POD, yang bertanggung jawab untuk mendetoksifikasi H 2 O 2 . Dengan meningkatkan pertahanan antioksidan utama ini, melatonin secara efektif mengurangi stres oksidatif, menjaga integritas sel, dan meningkatkan ketahanan tanaman terhadap kekurangan Fe.
Penambahan PAG membalikkan efek menguntungkan melatonin pada aktivitas enzim antioksidan. Dibandingkan dengan tanaman yang diberi melatonin, penambahan PAG menyebabkan penurunan substansial dalam aktivitas CAT dan POD dan mengurangi aktivitas SOD, sehingga memperburuk stres oksidatif. Hal ini menunjukkan bahwa kemampuan melatonin untuk meningkatkan pertahanan antioksidan terkait erat dengan sintesis H 2 S yang dimediasi L-DES, dan penghambatan jalur ini membahayakan efek perlindungan melatonin.
Menariknya, penggunaan NaHS, donor H 2 S eksogen, bersama melatonin dan PAG, memulihkan sebagian aktivitas enzim antioksidan. Dibandingkan dengan tanaman yang kekurangan Fe yang tidak diobati, aktivitas SOD meningkat sebesar 20%, aktivitas CAT meningkat sebesar 23%, dan aktivitas POD meningkat sebesar 60%. Hasil ini menyoroti bahwa suplementasi NaHS mengompensasi penghambatan sintesis H 2 S endogen yang disebabkan oleh PAG, memulihkan efek positif melatonin pada sistem pertahanan antioksidan.
3.6 Modulasi Kadar H 2 S dan Aktivitas L-DES sebagai Respon terhadap Melatonin dan PAG pada Defisiensi Zat Besi
Pada tanaman yang kekurangan zat besi, kadar H 2 S meningkat secara signifikan, lebih dari dua kali lipat dibandingkan dengan kontrol yang cukup Fe, disertai dengan peningkatan aktivitas L-DES yang nyata. Hal ini menunjukkan peningkatan regulasi jalur L-DES sebagai bagian dari respons tanaman terhadap kekurangan Fe (Gambar 5A,B ).
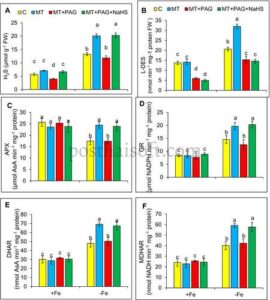
Namun, penggunaan PAG bersamaan dengan melatonin secara signifikan mengurangi kadar H 2 S hingga 40% dibandingkan dengan tanaman yang diberi melatonin, sementara aktivitas L-DES juga menurun. Hal ini menegaskan efektivitas PAG dalam menghambat aktivitas L-DES dan menyoroti ketergantungan produksi H 2 S yang diinduksi melatonin pada jalur L-DES.
Menariknya, penambahan NaHS, donor H 2 S eksogen, pada perlakuan melatonin + PAG memulihkan kadar H 2 S ke kadar yang sebanding dengan yang diamati pada tanaman yang diobati dengan melatonin saja. Akan tetapi, aktivitas L-DES tetap tidak berubah, yang selanjutnya menegaskan bahwa NaHS melewati penghambatan L-DES dengan secara langsung melengkapi H 2 S. Pemulihan kadar H 2 S ini secara efektif mendukung peran melatonin dalam mengurangi stres oksidatif dalam kondisi kekurangan Fe, bahkan dengan penghambatan L-DES.
3.7 Melatonin Meningkatkan Dinamika Redoks Askorbat dan Glutathione pada Kekurangan Zat Besi
Kekurangan zat besi mengganggu siklus AsA-GSH pada tanaman stroberi, mengubah aktivitas enzim utama dan kadar metabolit redoks. Dalam kondisi kekurangan Fe, aktivitas APX menurun hingga 32%, sementara aktivitas GR, DHAR, dan MDHAR meningkat masing-masing sebesar 72%, 66%, dan 58% (Gambar 5C–F ). Perubahan ini mencerminkan respons adaptif untuk mengurangi stres oksidatif dan mendetoksifikasi ROS. Bersamaan dengan itu, kadar AsA dan DHA hampir dua kali lipat, menghasilkan peningkatan rasio AsA/DHA sebesar 72%. Sementara itu, kadar total GSH dan GSSG meningkat, menyebabkan penurunan rasio GSH/GSSG sebesar 23,8%, yang menunjukkan ketidakseimbangan oksidatif (Gambar 6A–F ).
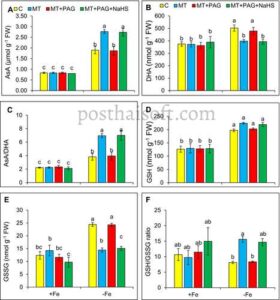
Namun, efek perlindungan melatonin berkurang secara signifikan dengan adanya PAG, karena aktivitas APX, GR, DHAR, dan MDHAR masing-masing menurun sebesar 29%, 36%, 30%, dan 29%. Rasio AsA/DHA menurun menjadi 3,95, sedangkan rasio GSH/GSSG hanya membaik sedikit, yang menunjukkan hilangnya sebagian keseimbangan redoks.
Penambahan NaHS bersama melatonin dan PAG memulihkan sebagian parameter antioksidan ini. Aktivitas APX, GR, DHAR, dan MDHAR meningkat masing-masing sebesar 38%, 62%, 34%, dan 36%, dan kadar AsA meningkat sementara kadar DHA menurun, sehingga meningkatkan rasio AsA/DHA. Kadar GSH tetap stabil, dan kadar GSSG menurun, sehingga meningkatkan rasio GSH/GSSG. Hasil ini menunjukkan bahwa H 2 S, yang diproduksi melalui L-DES, memainkan peran penting dalam memediasi mekanisme antioksidan melatonin dalam kondisi kekurangan Fe.
3.8 Aktivitas Metilglikosal dan Glioksalase dalam Respon terhadap Defisiensi Zat Besi dan Pengobatannya
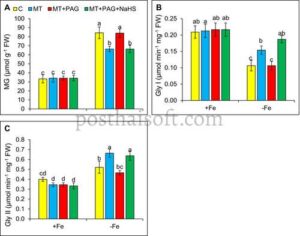
Pada defisiensi besi, level metilglioksal (MG) meningkat secara signifikan, naik sebesar 153% dibandingkan dengan tanaman yang cukup besi (Gambar 7A ). Sebaliknya, aktivitas glioksalase I (Gly-I) menurun, sementara aktivitas glioksalase II (Gly-II) meningkat, yang menunjukkan pergeseran metabolisme MG sebagai respons terhadap defisiensi besi (Gambar 7B,C ). Aplikasi melatonin pada defisiensi besi mengurangi level MG, penurunan sebesar 21% dibandingkan dengan tanaman yang tidak diobati defisiensi besi. Aktivitas Gly-I tetap stabil, sementara aktivitas Gly-II meningkat, yang menunjukkan bahwa melatonin meningkatkan konversi MG, yang mengarah pada peningkatan
aktivitas Gly-II.
4 Diskusi
4.1 Pengaturan Parameter Terkait Fotosintesis yang Diinduksi Melatonin pada Kekurangan Zat Besi
Studi ini meneliti bagaimana aktivitas L-DES dan sintesis H 2 S berkontribusi terhadap toleransi yang diinduksi melatonin terhadap stres defisiensi Fe pada tanaman stroberi. Hipotesis kami adalah bahwa aktivitas L-DES dan sintesis H 2 S meningkatkan efektivitas melatonin dalam mengurangi defisiensi Fe dengan mengurangi kerusakan oksidatif dan meningkatkan kinerja fisiologis. Sementara kemanjuran melatonin dalam mengurangi stres defisiensi Fe telah dibuktikan pada tanaman seperti Arabidopsis thaliana (Zhou et al. 2016 ), apel ( Malus domestica ) (Zheng et al. 2021 ), dan persik ( Prunus persica ) (Lin et al. 2023 ), mekanisme dasar yang melibatkan L-DES dan H 2 S masih belum sepenuhnya dipahami.
Jalur L-DES telah ditetapkan sebagai rute utama untuk sintesis H 2 S pada tanaman (Guo et al. 2017 ). Stres defisiensi besi, yang sering dikaitkan dengan pertumbuhan tanaman terhambat dan akumulasi H 2 O 2 berlebihan , telah dilaporkan pada spesies seperti Sulla carnosa (Jelali et al. 2021 ). Konsisten dengan temuan ini, penelitian ini menemukan bahwa defisiensi Fe menyebabkan kandungan klorofil tertekan dan kadar H 2 O 2 meningkat pada tanaman stroberi. Namun, aplikasi melatonin secara efektif mengurangi efek buruk ini dengan meningkatkan kandungan klorofil dan mengurangi konsentrasi H 2 O 2 , yang menggarisbawahi perannya sebagai modulator stres oksidatif yang kuat.
Hasil penelitian ini menunjukkan bahwa aktivitas L-DES dan produksi H 2 S merupakan elemen penting dari mekanisme toleransi stres yang diinduksi melatonin. Secara khusus, sintesis H 2 S tampaknya penting untuk mengurangi kerusakan oksidatif dalam kondisi kekurangan Fe, sebagaimana dibuktikan oleh berkurangnya efek melatonin setelah penghambatan sintesis H 2 S dengan PAG. Penghambatan ini tidak hanya membalikkan efek positif melatonin pada kandungan klorofil dan nutrisi Fe tetapi juga menggarisbawahi peran penting H 2 S yang dimediasi L-DES dalam mendukung ketahanan tanaman terhadap stres kekurangan Fe. Temuan ini mendukung penelitian sebelumnya yang menyoroti H 2 S sebagai pemain kunci dalam mitigasi stres yang dimediasi melatonin. Misalnya, H 2 S eksogen telah terbukti meningkatkan kandungan klorofil dan penyerapan Fe pada jagung yang kekurangan Fe ( Zea mays ) (Chen et al. 2015 ), stroberi ( Fragaria × ananassa ) (Kaya dan Ashraf 2019 ), dan kedelai ( Glycine max ) (He et al. 2023 ).
Selain efeknya pada stres oksidatif, melatonin secara signifikan meningkatkan efisiensi fotosintesis, seperti yang ditunjukkan oleh peningkatan rasio Fv/Fm di bawah stres defisiensi Fe. Peningkatan ini kemungkinan berasal dari kemampuan melatonin untuk meningkatkan transkripsi gen terkait fotosintesis seperti PsbA , PsbB , PsbC , PsbD , dan PsbO , yang mengkode protein kunci fotosistem II (PSII). Peran melatonin dalam mempromosikan ekspresi gen-gen ini di bawah stres telah dilaporkan pada tomat yang stres natrium-basa (Liu et al. 2015 ), tomat yang stres suhu tinggi (Jahan et al. 2021 ), dan paprika yang stres dingin (Altaf, Shu, et al. 2022 ).
Secara keseluruhan, temuan penelitian ini menunjukkan bahwa sintesis H 2 S yang dimediasi L-DES merupakan jalur penting dalam penanggulangan stres defisiensi Fe yang diinduksi melatonin. Dengan mengurangi kerusakan oksidatif dan meningkatkan efisiensi fotosintesis, melatonin memainkan peran ganda dalam meningkatkan ketahanan tanaman di bawah tekanan nutrisi.
4.2 Peran Melatonin dalam Pemanfaatan Zat Besi, Sinyal H 2 S, dan Aktivitas L-DES
Melatonin telah dikenal luas karena kapasitasnya untuk meningkatkan pemanfaatan Fe pada tanaman, terutama dalam kondisi stres. Peningkatan ini diperkirakan terjadi melalui mekanisme yang melibatkan reaktivasi dan pemanfaatan kembali zat besi, serta modulasi homeostasis Fe (Lin et al. 2023 ). Aspek penting dari peran melatonin dalam metabolisme Fe adalah interaksinya dengan pensinyalan H 2 S, yang mengaktifkan sistem pertahanan antioksidan dan memfasilitasi penyerapan dan asimilasi Fe (Kaya et al. 2020 ). Penelitian sebelumnya telah menyoroti kemampuan melatonin untuk mengatur gen yang terlibat dalam penyerapan zat besi, termasuk yang mempromosikan reduksi zat besi ferri (Fe 3+ ) menjadi bentuk ferro yang lebih mudah diserap (Fe 2+ ) melalui peningkatan aktivitas reduktase Fe (Zhou et al. 2016 ; Ahammed et al. 2020 ).
Bahasa Indonesia: Selain efek langsungnya pada metabolisme Fe, melatonin memengaruhi jalur metabolisme sulfur dengan menstimulasi produksi H 2 S melalui peningkatan aktivitas L-DES (Mukherjee dan Bhatla 2021 ). Studi ini memberikan bukti lebih lanjut yang mendukung peran penting H 2 S, yang dimediasi oleh aktivitas L-DES, dalam perbaikan yang diinduksi melatonin dalam metabolisme Fe. Penghambatan aktivitas L-DES oleh PAG, yang menekan sintesis H 2 S, membalikkan efek menguntungkan melatonin pada ketersediaan dan metabolisme Fe. Pengamatan ini menggarisbawahi pentingnya H 2 S sebagai molekul pensinyalan dalam memfasilitasi reduksi Fe 3+ menjadi Fe 2+ , sehingga meningkatkan bioavailabilitas Fe (Chen et al. 2020 ).
Menariknya, aplikasi H 2 S eksogen melalui aplikasi NaHS memulihkan efek menguntungkan melatonin pada metabolisme Fe, bahkan di bawah penghambatan L-DES. Temuan ini menunjukkan bahwa H 2 S berfungsi hilir L-DES dalam jalur yang dimediasi melatonin dan menyoroti hubungan silang antara melatonin, aktivitas L-DES, H 2 S, dan metabolisme Fe sebagai penentu penting toleransi tanaman terhadap defisiensi Fe (Chen et al. 2015 , 2020 ). Mengganggu jalur ini meniadakan efek perlindungan melatonin, menunjukkan bahwa sinergi antara melatonin dan H 2 S sangat diperlukan untuk menjaga homeostasis Fe dalam kondisi kekurangan nutrisi.
4.3 Melatonin Mengurangi Stres Oksidatif Akibat Kekurangan Zat Besi dengan Meningkatkan Aktivitas Antioksidan
Kekurangan zat besi merupakan penyebab stres oksidatif pada tanaman yang terdokumentasi dengan baik, terutama karena akumulasi ROS yang berlebihan. ROS ini dapat menimbulkan kerusakan seluler yang signifikan, termasuk peroksidasi lipid membran, denaturasi protein, dan gangguan fungsi seluler (Rahman et al. 2021 ). Melatonin, antioksidan yang kuat, dikenal luas karena kemampuannya untuk mengurangi stres oksidatif dengan meningkatkan mekanisme pertahanan antioksidan tanaman. Efek perlindungannya telah ditunjukkan dalam berbagai kondisi stres abiotik, termasuk kekeringan, salinitas (Siddiqui et al. 2021 ; Altaf, Shahid, et al. 2022 ; Mukherjee et al. 2023 ), dan kekurangan zat besi (Ahammed et al. 2020 ).
Dalam penelitian ini, fungsi antioksidan melatonin didukung lebih lanjut oleh regulasinya terhadap sintesis H 2 S melalui aktivitas L-DES. Penghambatan jalur ini oleh PAG mengurangi efek melatonin dalam meredakan stres, yang menekankan peran penting produksi H 2 S yang dimediasi L-DES dalam meningkatkan toleransi yang diinduksi melatonin. Konsisten dengan temuan pada mentimun di bawah tekanan garam (Sun et al. 2021 ), produksi H 2 S yang diinduksi melatonin bertindak sebagai mediator penting ketahanan stres, yang memperkuat perannya dalam mengurangi stres oksidatif di bawah kekurangan zat besi.
Kemampuan melatonin untuk memulihkan aktivitas enzim antioksidan utama, seperti SOD, CAT, dan POD, konsisten dengan temuan sebelumnya (Ahammed et al. 2020 ; Kaya et al. 2020 ). Enzim-enzim ini merupakan bagian penting dalam menetralkan ROS dan menjaga homeostasis redoks seluler. Dalam kondisi kekurangan zat besi, aktivitas enzim yang bergantung pada zat besi seperti CAT dan POD biasanya menurun karena berkurangnya ketersediaan Fe sebagai kofaktor. Namun, melatonin menangkal penurunan ini, melindungi tanaman dari kerusakan oksidatif dan menjaga keseimbangan redoks (Karumannil et al. 2023 ).
Lebih jauh lagi, melatonin dapat secara tidak langsung meningkatkan aktivitas Fe-reduktase dengan membatasi kerusakan sel yang disebabkan oleh ROS, sehingga meningkatkan ketersediaan dan penggunaan Fe dalam kondisi defisiensi (Huang et al. 2022 ). Pembalikan efek perlindungan melatonin setelah penghambatan L-DES, dan pemulihannya selanjutnya dengan aplikasi H 2 S eksogen melalui NaHS, menyoroti interaksi sinergis antara melatonin dan H 2 S dalam memperkuat sistem antioksidan tanaman. Interaksi ini sangat penting untuk meningkatkan ketahanan terhadap stres, sebagaimana dibuktikan oleh temuan serupa pada tanaman lain (Sun et al. 2021 ; Ghorbani et al. 2024 ).
4.4 Dinamika Redoks Askorbat dan Glutathione pada Pengobatan Defisiensi Zat Besi dan Melatonin
Stres defisiensi besi pada tanaman stroberi menyebabkan penekanan signifikan aktivitas APX, enzim kunci dalam siklus AsA-GSH. Penekanan ini dikaitkan dengan akumulasi asam askorbat (AsA), karena berkurangnya konsumsi AsA membatasi perannya dalam detoksifikasi ROS (Akram et al. 2017 ; Zha et al. 2020 ). Kadar AsA yang meningkat di bawah stres defisiensi besi berkontribusi pada pengurangan besi rizosfer, memfasilitasi penyerapan Fe, seperti yang diamati sebelumnya pada tanaman kacang polong (Grillet et al. 2014 ). Lebih jauh lagi, peningkatan rasio AsA/dehidroaskorbat (DHA) yang disebabkan oleh stres pada tanaman stroberi menunjukkan respons redoks spesifik terhadap defisiensi besi, berbeda dengan rasio yang berkurang yang dilaporkan pada tanaman lain di bawah berbagai kondisi stres (Singh et al. 2015 ). Temuan ini menunjukkan bahwa DHA memainkan peran penting dalam mengatur respons tanaman terhadap kerusakan oksidatif yang disebabkan oleh logam dan menjaga homeostasis redoks (Kaur dan Goyal 2022 ).
Perlakuan melatonin selanjutnya meningkatkan kadar AsA dan menstabilkan rasio AsA/DHA, yang menunjukkan perannya dalam meningkatkan kapasitas penyangga redoks tanaman (Zhang et al. 2017 ). Namun, PAG, penghambat L-DES, menghilangkan efek perlindungan melatonin pada dinamika AsA dengan menekan sintesis H 2 S. Aplikasi H 2 S eksogen melalui NaHS memulihkan efek positif melatonin, yang menyoroti peran penting produksi H 2 S yang dimediasi L-DES dalam mengatur keseimbangan AsA/DHA.
Demikian pula, stres defisiensi besi meningkatkan kadar glutathione tereduksi (GSH) dan bentuk teroksidasinya (GSSG) pada tanaman stroberi, yang menyebabkan penurunan rasio GSH/GSSG. Respons ini kontras dengan pengamatan pada bit gula, di mana GSH dan rasio GSH/GSSG tetap tidak berubah di bawah defisiensi besi (Zaharieva dan Abadía 2003 ). Praperlakuan melatonin berhasil memulihkan kadar GSH dan menormalkan rasio GSH/GSSG, konsisten dengan efeknya pada tomat di bawah stres kekeringan (Altaf, Shahid, et al. 2022 ) dan mentimun di bawah stres salinitas (Wang et al. 2016 ). Penghambatan L-DES oleh PAG mengganggu homeostasis redoks yang diinduksi melatonin ini, sementara suplementasi NaHS memulihkan kadar GSH dan keseimbangan GSH/GSSG.
Peran NaHS eksogen dalam meningkatkan dinamika GSH didukung oleh penelitian sebelumnya yang menunjukkan bahwa NaHS meningkatkan aktivitas GSH pada gandum di bawah tekanan panas (Iqbal et al. 2021 ) dan pada tanaman kacang yang terpapar toksisitas arsenik (Alsahli et al. 2021 ). Temuan-temuan ini secara kolektif menekankan pentingnya produksi H 2 S yang dimediasi L-DES dalam mengatur dinamika siklus AsA-GSH dan menjaga keseimbangan redoks di bawah tekanan defisiensi besi. Dengan memodulasi status redoks, melatonin dan interaksinya dengan H 2 S bertindak sebagai mediator utama ketahanan tanaman terhadap stres nutrisi.
4.5 Peningkatan Enzim Siklus AsA-GSH yang Dimediasi Melatonin sebagai Respon terhadap Defisiensi Zat Besi
Tumbuhan meningkatkan pertahanan antioksidannya dengan mensintesis AsA dan GSH dalam kondisi stres, yang keduanya penting untuk membersihkan ROS dan menjaga keseimbangan redoks (Jiang et al. 2022 ; Peng et al. 2022 ). Pengaturan melatonin terhadap enzim utama dalam jalur AsA-GSH telah banyak dilaporkan, yang menggarisbawahi perannya dalam meningkatkan ketahanan tanaman terhadap berbagai stres abiotik.
Dalam penelitian ini, defisiensi zat besi menyebabkan penurunan aktivitas APX yang nyata pada tanaman stroberi, konsisten dengan pengamatan pada alfalfa (Rahman et al. 2021 ) dan padi (Zhang, Xue, et al. 2022 ). APX sangat penting untuk mendetoksifikasi H 2 O 2 dalam jalur AsA-GSH, dan penekanannya di bawah tekanan defisiensi zat besi memperburuk akumulasi ROS. Namun, praperlakuan melatonin secara signifikan memulihkan aktivitas APX, konsisten dengan temuan sebelumnya pada tomat di bawah tekanan kekeringan (Altaf, Shahid, et al. 2022 ).
Demikian pula, glutathione reductase (GR), yang meregenerasi GSH dari bentuk teroksidasinya (GSSG), menunjukkan peningkatan aktivitas di bawah tekanan kekurangan zat besi pada tanaman stroberi, konsisten dengan laporan sebelumnya pada spesies lain (Santos et al. 2019 ). Melatonin selanjutnya meningkatkan aktivitas GR, meningkatkan efisiensi siklus AsA-GSH dan mempromosikan homeostasis zat besi melalui detoksifikasi ROS yang ditingkatkan. Namun, efek menguntungkan melatonin pada aktivitas APX dan GR dihapuskan ketika PAG, penghambat L-DES, diterapkan. Ini menyoroti peran penting produksi H 2 S yang dimediasi L-DES dalam toleransi stres yang diinduksi melatonin.
Sejauh pengetahuan kami, penelitian ini memberikan bukti pertama bahwa melatonin mengaktifkan L-DES untuk menghasilkan H 2 S, yang pada gilirannya meningkatkan aktivitas APX dan GR di bawah tekanan kekurangan zat besi. Temuan ini menggarisbawahi peran saling terkait dari melatonin, L-DES, dan H 2 S dalam mengatur aktivitas enzim antioksidan dan mempertahankan homeostasis redoks.
Selain itu, enzim lain dalam siklus AsA-GSH, seperti DHAR dan MDHAR, meningkat di bawah tekanan defisiensi zat besi dan lebih ditingkatkan lagi dengan pengobatan melatonin. Enzim-enzim ini memainkan peran penting dalam mendaur ulang AsA, mengurangi H 2 O 2 , dan mempertahankan keseimbangan redoks seluler, seperti yang diamati dalam penelitian sebelumnya (Hasanuzzaman et al. 2019 ; Namdjoyan dan Mohammadian 2024 ). Kemampuan melatonin untuk meningkatkan aktivitas DHAR dan MDHAR menyoroti peran komprehensifnya dalam memodulasi siklus AsA-GSH dan meningkatkan ketahanan tanaman terhadap defisiensi nutrisi.
4.6 Akumulasi Metilglikosal dan Perubahan Jalur Glioksalase
Metilglioksal (MG), senyawa α-dikarbonil reaktif, terakumulasi dalam sel tanaman dalam berbagai kondisi stres, termasuk kekurangan zat besi. Akumulasi ini terkait erat dengan stres oksidatif, karena MG mendorong pembentukan ROS dan mengganggu sistem antioksidan, yang menyebabkan efek sitotoksik (Cai et al. 2019 ). Kadar MG yang tinggi mengganggu homeostasis seluler, merusak fungsi metabolisme, dan memperburuk kerusakan oksidatif, yang selanjutnya menghambat pertumbuhan dan perkembangan tanaman, terutama dalam kondisi stres nutrisi seperti kekurangan zat besi (Alhammad et al. 2023 ).
Tumbuhan menangkal toksisitas MG melalui sistem glioksalase, jalur detoksifikasi penting yang terdiri dari glioksalase I (Gly-I) dan glioksalase II (Gly-II). Enzim-enzim ini memfasilitasi konversi MG menjadi senyawa yang kurang berbahaya, seperti D-laktat, dengan menggunakan glutathione (GSH) sebagai kofaktor. Aktivitas glioksalase yang berubah pada kondisi kekurangan zat besi menunjukkan bahwa tumbuhan secara aktif berupaya mengurangi toksisitas MG sambil mengatasi proses metabolisme yang terganggu (Li 2016 ; Garai et al. 2021 ).
Kekurangan zat besi biasanya mengakibatkan peningkatan kadar MG karena gangguan regulasi metabolik dan berkurangnya pertahanan antioksidan. Namun, aplikasi melatonin eksogen telah terbukti meringankan sitotoksisitas yang diinduksi MG dengan memodulasi aktivitas glioksalase. Melatonin mengurangi konsentrasi MG dan meningkatkan aktivitas Gly-I dan Gly-II, sebagaimana dibuktikan dalam penelitian terbaru (Altaf et al. 2024 ; Imran et al. 2024 ). Dalam penelitian ini, pengobatan melatonin secara signifikan meningkatkan aktivitas Gly-I dan Gly-II, yang menunjukkan peralihan ke arah detoksifikasi MG yang lebih efisien dan pemulihan keseimbangan metabolik. Hal ini sejalan dengan temuan sebelumnya yang menunjukkan bahwa regulasi aktivitas glioksalase yang dimediasi melatonin berkontribusi pada pengurangan stres oksidatif dan peningkatan toleransi stres pada tanaman.
5 Kesimpulan
Studi ini menggarisbawahi peran penting melatonin dalam mengurangi stres akibat kekurangan zat besi pada tanaman stroberi, terutama melalui aktivasi L-DES dan produksi H 2 S berikutnya. Perlakuan melatonin secara signifikan meningkatkan bioavailabilitas Fe dan kandungan klorofil, sehingga menangkal efek merugikan dari kekurangan Fe. Aktivasi enzim antioksidan seperti SOD dan CAT dalam perlakuan melatonin semakin menyoroti perannya dalam memperkuat sistem pertahanan antioksidan terhadap stres oksidatif.
Selain itu, modulasi dinamika redoks askorbat-glutathione, yang ditandai dengan peningkatan kadar askorbat dan glutathione serta rasio redoks yang dipulihkan, memainkan peran penting dalam menjaga keseimbangan redoks seluler. Melatonin juga memfasilitasi detoksifikasi metilglioksal melalui jalur glioksalase, yang menunjukkan perannya yang beragam dalam meningkatkan ketahanan metabolik pada defisiensi zat besi. Temuan ini menekankan interaksi sinergis antara melatonin, L-DES, dan H 2 S dalam mempromosikan homeostasis Fe, yang menyediakan kerangka mekanistik untuk menggunakan melatonin sebagai biostimulan guna meningkatkan ketahanan tanaman terhadap defisiensi Fe.
6 Prospek Masa Depan
Penelitian di masa mendatang harus difokuskan pada pengungkapan mekanisme molekuler yang mendasari interaksi antara melatonin dan L-DES, khususnya jalur regulasi yang mengatur sintesis H 2 S dan dampaknya pada mobilisasi dan penyerapan Fe. Investigasi terhadap modifikasi transkripsi dan pascatranslasi enzim utama yang terlibat dalam jalur ini akan memberikan wawasan yang lebih mendalam tentang fungsionalitasnya dalam berbagai kondisi stres.
Perluasan cakupan penelitian melatonin agar mencakup spesies tanaman dan tekanan lingkungan yang lebih luas, seperti salinitas, kekeringan, dan keracunan logam berat, dapat mengungkap penerapannya yang lebih luas dalam pengelolaan tanaman. Studi lapangan jangka panjang yang menilai dampak melatonin terhadap hasil, kualitas buah, dan efisiensi penggunaan nutrisi sangat penting untuk menentukan kelayakan praktisnya.
Selain itu, penelitian tentang integrasi aplikasi melatonin dengan praktik agronomi lainnya, seperti amandemen tanah dan strategi pengelolaan nutrisi, dapat meningkatkan produktivitas tanaman dan ketahanan pangan, terutama di wilayah yang rentan terhadap defisiensi Fe.