
ABSTRAK
Tujuan
Penggembalaan berlebihan oleh populasi rusa yang berlebihan merusak komunitas tumbuhan asli di sebagian besar wilayah beriklim sedang dan boreal utara. Di wilayah yang dampak rusanya sangat parah, pemagaran terkadang digunakan sebagai tindakan konservasi untuk melindungi spesies dan ekosistem yang langka dan terancam. Di sini kami meneliti bagaimana komunitas tumbuhan vaskular hutan telah terpengaruh oleh pengecualian rusa sika selama 21 tahun dengan pemagaran di area seluas 900 ha.
Lokasi
Taman Nasional Nikko, prefektur Tochigi, Jepang.
Metode
Kami mensurvei tutupan tumbuhan vaskular di 240 plot dan mengukur kerapatan hutan di kedua sisi pagar pada tegakan yang didominasi oleh larch ( Larix kaempferi ) dan oak ( Quercus crispula ).
Hasil
Kekayaan spesies tumbuhan berpembuluh ditemukan lebih rendah di kawasan hutan yang dilindungi oleh pagar. Pada saat yang sama, bambu kerdil, yang sering kali merupakan komponen tumbuhan bawah yang paling melimpah di hutan terbuka Jepang, lebih tinggi dan lebih rapat di dalam pagar tempat rusa telah disingkirkan. Selain itu, kurangnya regenerasi pohon alami di dalam pagar, yang kemungkinan besar disebabkan oleh persaingan bambu kerdil yang ketat, juga terlihat jelas. Hal ini menunjukkan bahwa pengusiran rusa telah menguntungkan bambu kerdil sekaligus menghambat persaingan spesies lain, sehingga menurunkan kekayaan tumbuhan berpembuluh. Komunitas tumbuhan berpembuluh di luar pagar tampaknya dibentuk dengan kuat oleh rusa sika yang melimpah; beberapa spesies yang ditemukan di sana diketahui tidak enak dimakan atau toleran terhadap penggembalaan, dan hanya ada sedikit bibit pohon.
Kesimpulan
Hasil ini menunjukkan bahwa pagar untuk melindungi flora hutan dari penggembalaan berlebihan dapat menimbulkan konsekuensi yang tidak diinginkan karena ada risiko spesies kompetitif dapat mendominasi komunitas tumbuhan bawah. Alih-alih pagar, pengelolaan hewan buruan yang lebih baik untuk mempertahankan populasi rusa dalam jumlah sedang mungkin merupakan strategi yang lebih baik untuk konservasi flora hutan dalam skala besar dan untuk mendorong regenerasi pohon alami.
1 Pendahuluan
Populasi ungulata yang berlebihan mengubah komposisi dan struktur vegetasi hutan di sebagian besar wilayah utara beriklim sedang dan boreal (Rooney dan Waller 2003 ; Miyaki dan Kaji 2009 ; Kishimoto et al. 2010 ; Petersson, Bergstedt, et al. 2019 ; Reed et al. 2022 ). Penggembalaan rusa yang intensif bahkan dapat menyebabkan tanah longsor dengan menghilangkan vegetasi lantai hutan dan melonggarkan cengkeraman sistem akar pada tanah (Wakahara et al. 2008 ). Pencarian makanan selektif rusa juga memengaruhi keanekaragaman bunga dengan mengubah hubungan kompetitif di antara spesies tanaman vaskular (Côté et al. 2004 ). Dampak komunitas tanaman dari penggembalaan rusa yang berlebihan dapat berlangsung lama atau bahkan tidak dapat dipulihkan ketika spesies tersebut punah dari bank benih (DiTommaso et al. 2014 ; Nuttle et al. 2014 ; Tamura 2016 ).
Wilayah Jepang yang dihambat oleh rusa sika ( Cervus nippon ) telah meningkat dari 25% menjadi 70% antara tahun 1978 dan 2018, dan perkiraan populasi rusa di Jepang di luar Hokkaido telah tumbuh dari 250.000 pada tahun 1989 menjadi 2.180.000 pada tahun 2020 (MOE 2021 ; Anonymus 2022 ). Beberapa faktor kemungkinan berada di balik pertumbuhan yang cepat ini. Ketika serigala abu-abu Jepang ( Canis lupus hodophilax ) punah pada tahun 1905 karena perburuan, satu-satunya predator alami rusa sika menghilang (Walker 2005 ). Selama waktu itu dan sepanjang paruh pertama abad ke-20, jumlah rusa sika di Jepang sangat rendah (Uno et al. 2022 ). Dalam upaya untuk meningkatkan populasi rusa, perburuan rusa sika betina dilarang sejak tahun 1963 (Oka et al. 2022 ). Pada saat yang sama, urbanisasi yang cepat menyebabkan lahan pertanian dan kebun buah yang terbengkalai luas, yang menyediakan habitat rusa yang baru dan sesuai. Ketika larangan berburu rusa betina dicabut secara nasional pada tahun 2007, populasi rusa telah mencapai tingkat yang menyebabkan kerusakan signifikan pada pertanian dan kehutanan (Oka et al. 2022 ). Baru-baru ini, perubahan iklim telah membuat hujan salju lebat dan suhu musim dingin yang sangat rendah secara berkala menjadi lebih jarang terjadi, sehingga meningkatkan kelangsungan hidup rusa muda, yang memicu perluasan populasi lebih lanjut (Ueno et al. 2018 ).
Di daerah hutan beriklim sedang di Jepang, makanan pokok rusa sika adalah beberapa spesies bambu kerdil dari genus Neosasamorpha , Pleioblastus , Pseudosasa , Sasa , dan Sasamorpha (Campos-Arceiz dan Takatsuki 2005 ; Murata et al. 2009 ; Kobayashi 2015 ; Kagamiuchi dan Takatsuki 2020 ; Takatsuki 2022 ). Mereka tersebar luas di zona beriklim sedang di Asia timur, di mana mereka sering membentuk lapisan bawah hutan terbuka hingga semi terbuka (Kobayashi 2015 ). Setelah gangguan, misalnya, penebangan hutan dan kebakaran, bambu kerdil dapat membentuk petak klonal padat yang menutupi area yang luas (Oshima 1961 ). Jika tidak terganggu, tegakan bambu kerdil murni dapat bertahan tanpa batas waktu, terutama di hutan terbuka dan lingkungan pegunungan (Oshima 1961 ). Di sini, mereka mendominasi komunitas tumbuhan tingkat bawah dan bahkan dapat menyingkirkan spesies tingkat bawah lainnya (Fukamachi et al. 1996 ; Itô dan Hino 2005 ; Kudo et al. 2011 ; Černý et al. 2013 ). Di area dengan tutupan salju musim dingin, rusa sika terus memakan bambu kerdil selama masih dapat diakses. Namun, ketika salju menjadi terlalu tebal (sekitar 100 cm), rusa sika semakin banyak mencari makan di kulit kayu, merusak dan terkadang membunuh pohon (Minamino dan Akashi 2011 ). Selama musim panas dan di area tanpa tutupan salju, kebiasaan mencari makan rusa sika lebih bervariasi. Sementara pengupasan kulit kayu di musim panas juga telah diamati (Yokoyama et al. 1996 ), makanan mereka sering kali mencakup berbagai tumbuhan tingkat bawah selain bambu kerdil, serta ranting dan daun pohon (Yokoyama et al. 1996 ; Miyaki dan Kaji 2004 ; Iijima dan Nagaike 2015 ). Populasi rusa sika yang padat saat ini telah menyebabkan perubahan yang dilaporkan pada komunitas tumbuhan di berbagai wilayah Jepang (Yokoyama dan Shibata 1998 ; Kato dan Okuyama 2004 ; Murata et al. 2009 ; Uno et al. 2019 ). Kekhawatiran juga telah muncul mengenai potensi dampak jangka panjang dari penggembalaan rusa secara intensif, yang dapat menimbulkan dampak yang mendalam dan tidak dapat dipulihkan pada flora (Takatsuki 2009 ).
Komunitas lantai hutan sangat dipengaruhi oleh kondisi lokasi dan karakteristik pohon lantai hutan (Barbier et al. 2008 ; Petersson, Holmström, et al. 2019 ). Spesies pohon lantai hutan dan kerapatan pohon memengaruhi ketersediaan cahaya yang ditransmisikan ke komunitas lantai hutan (Lariviere et al. 2021 ; Petersson et al. 2021 ). Serasah dari pohon membentuk lapisan tanah, yang selanjutnya memengaruhi ketersediaan nutrisi dan kelembapan tanah, yang selanjutnya memengaruhi komposisi komunitas lantai hutan (Xiong dan Nilsson 1999 ).
Dalam penelitian ini, kami bermaksud untuk meneliti efek gabungan dari penggembalaan rusa sika, spesies pohon tajuk, dan kepadatan tegakan di suatu area Taman Nasional Nikko, Prefektur Tochigi, Jepang. Untuk pertama kalinya, dalam artikel ini kami meneliti bagaimana pengucilan rusa sika memengaruhi komunitas tumbuhan vaskular tajuk bawah di area yang luas yang terdiri dari berbagai jenis hutan. Dengan membandingkan vegetasi di dalam dan di luar kandang rusa seluas ~900 ha yang telah dipagari selama 21 tahun, bersama dengan informasi tentang spesies pohon dan kepadatan tegakan, kami berupaya untuk menjawab pertanyaan penelitian berikut:
Bagaimana flora tumbuhan vaskular hutan terpengaruh oleh pengecualian rusa sika?
Bagaimana pengecualian rusa memengaruhi kekayaan spesies dan komposisi komunitas tumbuhan vaskular hutan?
Bagaimana spesies pohon tajuk dan kerapatan hutan mempengaruhi kekayaan spesies tanaman vaskular di dalam dan luar pagar?
2 Metode
2.1 Wilayah Studi
Penelitian ini dilakukan pada tahun 2022 di Taman Nasional Nikko di Prefektur Tochigi, Jepang (36.77° LU, 139.43° BT, elevasi 1380–1470 m) di lembah dataran tinggi yang luas di daerah pegunungan (Gambar 1 ). Suhu rata-rata di daerah ini adalah 7,1°C, dan curah hujan rata-rata 2102 mm antara tahun 2002 dan 2011 (Tadakara et al. 2014 ). Batuan dasarnya mengandung silika, terutama terdiri dari berbagai bentuk granit dan riolit (Yamamoto et al. 2000 ). Tanahnya ditutupi oleh andosol haplik dan podzol (Oyama et al. 1969 ; Yamamoto et al. 2000 ). Danau Chuzenji dan Gunung Nantai adalah landmark menonjol di daerah ini, yang merupakan tujuan populer bagi wisatawan karena pemandangan hutan, danau, air terjun, lahan basah, dan pegunungan yang indah. Di atas Danau Chuzenji terletak dataran tinggi dan rawa Senjogahara, yang terbentuk oleh letusan Gunung Nantai sekitar 20.000 tahun yang lalu.
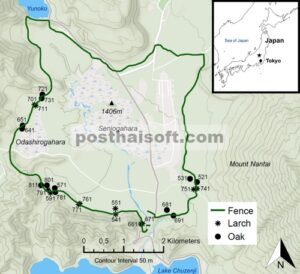
Area studi terletak di Taman Nasional Nikko (★), Jepang. Sepasang plot ditempatkan di setiap sisi pagar di 12 tegakan larch dan oak. Nomor tegakan tiga digit dapat digunakan untuk membandingkan kekayaan spesies tumbuhan berpembuluh dan tutupan di berbagai tegakan (lihat Lampiran S2 , Gambar S1 ). Sumber: OpenStreetMap.
Flora rawa Senjogahara mencakup tutupan Spiraea salicifolia yang melimpah . Rawa tersebut bergeser dari lahan basah menjadi hutan karena suksesi alami dan drainase buatan sebelumnya. Oleh karena itu, zona perbatasan antara rawa yang sebelumnya dikeringkan didominasi oleh tegakan semi-terbuka dari spesies pohon Malus toringo yang suksesi awal . Sebagian besar hutan di sekitar Senjogahara terdiri dari tegakan satu lapis larch Jepang ( Larix kaempferi , di bawah ini disebut sebagai “larch”) dan ek Mongolia ( Quercus crispula , sinonim. Quercus mongolica var. crispula , di bawah ini disebut “ek”). Tegakan tersebut mungkin tumbuh setelah area hutan tua yang luas ditebang setelah Perang Dunia II. Saat ini, hanya beberapa pohon larch dan ek yang tersisa dari generasi pohon yang lebih tua yang tersisa di antara tegakan yang usianya sama (Gambar 2 ). Spesies pohon lain yang umum tumbuh di kawasan tersebut adalah Abies homolepis , A. firma , Acer spp., Betula spp., Fagus crenata , Fraxinus mandshurica , Kalopanax septemlobus subsp. septemlobus , Pterocarya rhoifolia , Sambucus racemosa subsp. sieboldiana , dan Ulmus davidiana var. japonica .

Pagar rusa di area yang didominasi pohon ek. Area di sebelah kiri berada di dalam pagar, di sini didominasi oleh bambu kerdil. Di sebelah kanan, area di luar pagar memiliki semak belukar yang didominasi oleh Aster leiophyllus var. leiophyllus yang tidak enak dimakan . Di dekat bagian tengah gambar, sisa pohon ek yang lebih tua dapat terlihat. Gambar diambil di bagian tenggara area berpagar (Gambar 1 ). Foto: Lisa Petersson.
2.2 Pagar Rusa
Untuk melindungi flora langka di tanah rawa Senjogahara dan untuk mempromosikan regenerasi hutan di daerah sekitarnya, sebuah pagar dipasang pada tahun 2001 (Bansho dan Amemiya 2010 ). Pagar tersebut melingkupi area seluas 900 ha, termasuk tanah rawa Senjogahara, tanah rawa Odashirogahara yang lebih kecil, dan area hutan yang bersebelahan (Gambar 1 ). Pagar tersebut bertujuan untuk sepenuhnya mengecualikan rusa sika dari area dalam. Namun, terutama selama musim dingin, rusa kadang-kadang masuk ke dalam pagar (Bansho dan Amemiya 2010 ). Setelah upaya yang ditargetkan untuk menyingkirkan rusa yang mengganggu dimulai pada tahun 2005, jumlah rusa yang ada di dalam pagar telah menurun (Bansho dan Amemiya 2010 ). Namun, rusa kadang-kadang masih terlihat di dalam pagar (pengamatan pribadi), tetapi mereka secara aktif disingkirkan selama musim dingin sebagai pengendalian satwa liar yang berbahaya, dan akibatnya, herbivori rusa rendah.
2.3 Pemilihan Stand
Kami mengidentifikasi 24 tegakan hutan yang cocok untuk penelitian, masing-masing 12 tegakan larch dan oak, di bagian selatan area berpagar (Gambar 1 ). Tegakan tersebut dipilih sehingga replikat dengan spesies pohon dominan yang sama ditempatkan sedekat mungkin di sisi berlawanan dari pagar. Jika tidak ada replikat tegakan yang ditemukan dalam jarak 300 m, tegakan tersebut tidak digunakan. Kami juga memastikan bahwa pasangan tegakan hutan yang dipilih terletak pada ekosite yang sama, dengan umur pohon, kerapatan tegakan, topografi, dan sifat tanah yang sebanding.
Hanya tegakan larch atau oak di area barat daya yang disertakan dalam penelitian. Beberapa bagian timur dan barat area berpagar didominasi oleh pohon peluruh lainnya, sehingga tidak dapat digunakan. Bagian timur laut pagar didominasi oleh tegakan larch dan oak; namun, kami tidak dapat melihat tanda-tanda rusa merumput di luar pagar di area ini. Alasan mengapa rusa tidak mencari makan di sini tidak jelas. Hal ini mungkin disebabkan oleh topografi kipas aluvial yang curam dan Sungai Sakasa serta ngarai Misawa, yang dapat membatasi akses ke area di luar pagar.
2.4 Inventarisasi Plot
Inventarisasi tumbuhan vaskular dilakukan antara 21 Juli dan 20 September 2022. Sepuluh plot melingkar per tegakan dibuat, dengan total 240 plot melingkar (120 di dalam dan 120 di luar pagar). Setiap plot mencakup area seluas 10 m 2 dan dipusatkan pada pohon larch atau oak. Karena plot dibuat di sekitar pohon (bukan di ruang di antara pohon), hasilnya terutama mencerminkan vegetasi langsung di bawah tajuk daripada area nonpohon lainnya di tegakan. Dimulai dari pohon referensi tegakan di pusat tegakan, sembilan pohon hidup lainnya > 10 cm diameter pada 130 cm di atas tanah dipilih sebagai pusat plot tambahan. Ini dipilih sehingga dua atau tiga plot terletak di setiap arah mata angin (Gambar 3 ). Jarak minimum antara pohon pusat plot adalah 5 m, dan plot ditempatkan pada posisi pertama yang dapat diterima menurut arah mata angin dan jarak minimum antara pusat plot. Kadang-kadang, pohon tunggal atau kelompok kecil seperti Abies , Acer , Betula , atau Kalopanax ditemukan di dalam tegakan. Untuk menghindari potensi pengaruh dari pohon tajuk selain larch dan oak, petak hanya ditempatkan minimal 10 m dari pohon spesies lain dengan diameter batang > 30 cm.
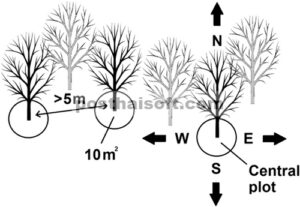
Tata letak pemilihan plot. Pertama, pohon referensi tegakan diidentifikasi di tengah setiap tegakan. Dari pohon referensi tegakan, dua atau tiga plot tambahan dipilih di masing-masing dari empat arah mata angin, dengan total 10 plot per tegakan. Semua plot dibuat dengan jarak minimal 5 m antara pohon-pohon di tengah plot.
Bahasa Indonesia: Di setiap plot, tinggi bambu kerdil tingkat bawah ditentukan dari rata-rata tinggi yang diukur dari lima titik acak per plot. Kami juga mengukur diameter dan mencatat spesies semua pohon di sekitarnya dalam radius 7 m dari pusat plot. Selain itu, kami mencatat arah kompas dan jarak ke pusat plot untuk setiap pohon yang diukur. Semua spesies tumbuhan vaskular tingkat bawah, termasuk pohon dan semak < 1,3 m, dicatat. Tutupan spesies yang berbeda diukur menggunakan versi modifikasi kelas tutupan Braun–Blanquet dengan tujuh kategori tutupan yang berbeda (Tabel 1 ). Dalam versi modifikasi ini, ada lebih banyak penekanan pada perbedaan tutupan antara kelas tutupan kecil. Perhatikan bahwa ketika menambahkan spesies yang berbeda yang tumbuh bersarang bersama di plot yang sama, tutupan total dapat melebihi 100%.
TABEL 1. Kelas Braun–Blanquet yang digunakan untuk mengukur tutupan tumbuhan vaskular.
Skala Braun–Blanquet Jangkauan cakupan (%) Digunakan dalam perhitungan (%)
R < 0,1 0,05 + > 0,1–1 0,55
1 > 1–5 3
2 > 5–25 15
3 > 25–50 37.5
4 > 50–75 62.5
5 > 75–100 87,5 atau 100 a
a Untuk bambu kerdil, yang jumlahnya sangat banyak, digunakan tutupan 100% untuk Kategori 5. Penggunaan salah satu angka tersebut menghasilkan hasil yang serupa.
Tumbuhan tertentu tidak dapat diidentifikasi spesiesnya dan malah dicatat pada tingkat genus atau famili dan kadang-kadang pada tingkat taksonomi atau kelompok fungsional yang lebih tinggi (misalnya, pakis, herba, tanaman merambat; lihat daftar spesies di Lampiran S1 ). Demi kenyamanan, kami mengelompokkan semua spesies bambu kerdil Sasa kurilensis , S. nipponica , S. palmata , dan Sasamorpha borealis yang sering membentuk area luas tempat berbagai spesies tumbuh bercampur. Tumbuhan ini secara kolektif dicatat sebagai kelompok Sasa spp. dalam daftar spesies. Karena kami jarang menemukan spesimen berbunga dari graminoid lain, tidak mungkin untuk membedakan antar spesies, dan mereka harus digabungkan sebagai Carex , Juncus , Luzula , dan Poaceae. Tumbuhan tidak berbunga lainnya yang dicatat pada tingkat genus termasuk Rosa , Rubus , dan Viola (tidak termasuk Viola tokubuchiana var. tokubuchiana ). Selain itu, karena tantangan dalam mengidentifikasi bibit pohon kecil di lapangan, kami mencatat semua spesies Acer bersama-sama kecuali A. rufinerve , yang dicatat secara terpisah. Pohon Betula platyphylla var. japonica dan B. ermanii dikelompokkan sebagai Betula , dan semua pohon dan semak Euonymus dikelompokkan bersama, yaitu E. alatus , E. macropterus , dan E. sieboldianus , sementara memperlakukan tanaman merambat E. fortunei sebagai entitas yang terpisah. Tata nama mengikuti daftar periksa Wamei versi 1.10 (Yamanouchi et al. 2022 ).
2.5 Indeks Pengaruh Pohon
Untuk meneliti bagaimana kekayaan spesies tumbuhan berpembuluh dapat dipengaruhi oleh kepadatan tegakan yang berbeda, kami menghitung indeks pengaruh pohon (TI). Indeks ini menggabungkan diameter pohon, arah mata angin, dan jarak dari pusat plot contoh ke pohon-pohon di sekitarnya sebagai berikut:
di mana ϴ i adalah arah kompas dari pusat plot ke pohon, C i adalah keliling pohon dan D i adalah jarak dari pohon pusat plot. Semua pohon dan semak > 1,3 m tinggi ( i ) dalam jarak 7 m (diperpanjang hingga 10 m ketika kurang dari empat pohon muncul) dari pusat plot dimasukkan dalam perhitungan indeks TI. Indeks ini adalah jumlah dari total luas dasar pohon ( C i 2 /4 π ) di sekitar pusat plot yang dibobot oleh dua faktor. Faktor pertama adalah jarak ( D i ) ke pohon yang pengaruhnya kita anggap berkurang dengan kuadrat jarak, analog dengan banyak hukum fisika. Yang kedua adalah arah kompas ( ϴ ) ke pohon. Istilah ((− cos ((
−180)/180) + 1) × 0,4 + 0,2) mengubah arah ini menjadi faktor yang berskala dari 1 untuk pohon di selatan pusat hingga 0,2 untuk pohon di utara, dan 0,6 untuk pohon di timur atau barat. Transformasi ini bertujuan untuk merepresentasikan kontribusi relatif pohon di berbagai arah terhadap naungan. Nilai TI berkisar dari 25,67 hingga 916,64 di berbagai plot, dengan rata-rata 291,51 ± SD 162,11. Sebelum melakukan analisis regresi, nilai dinormalisasi melalui penskalaan dengan pemusatan menggunakan fungsi scale() di R.
3 Metode Statistik
3.1 Tutupan dan Tinggi Bambu Kerdil dan Tumbuhan Berpembuluh
Perbedaan tutupan dan tinggi bambu kerdil, dan total tutupan tanaman berpembuluh di dalam dan di luar pagar dibandingkan pada tingkat tegakan menggunakan uji- t berpasangan . Tegakan yang berdekatan satu sama lain, tetapi pada sisi berlawanan dari pagar (Gambar 1 ) ditetapkan sebagai pasangan dalam analisis. Pada pengujian pertama, semua tegakan diuji bersama-sama. Pada pengujian kedua, variabel yang sama diuji secara terpisah untuk tegakan larch dan oak.
3.2 Model Kekayaan Spesies
Kekayaan spesies pada tingkat plot diuji menggunakan model campuran linier umum (GLMM) dengan distribusi Poisson menggunakan paket R “lme4” (Kuznetsova et al. 2017 ). Untuk memperhitungkan struktur hierarki data, dua efek acak ditetapkan dalam model. Pertama, karena setiap tegakan terdiri dari 10 plot, tegakan variabel juga ditetapkan sebagai efek acak dalam analisis. Kedua, dengan mempertimbangkan kedekatan antara tegakan di sisi berlawanan dari pagar, tegakan berpasangan ditetapkan sebagai efek acak.
Mengingat korelasi kuat antara tinggi bambu kerdil dan sisi pagar (yaitu, bambu kerdil secara konsisten lebih tinggi di dalam pagar daripada di luar; Gambar 4b ), model GLMM terpisah dibangun untuk menilai efek pagar dan tinggi dan penutup bambu kerdil. Model pertama bertujuan untuk mengevaluasi pengaruh tiga variabel: sisi pagar (biner), spesies pohon tajuk (biner), dan indeks pengaruh pohon (TI—kontinu). Dalam model lengkap pertama, kekayaan spesies diperiksa untuk interaksi antara spesies pohon, sisi pagar (di dalam atau di luar), dan TI. Namun, karena tidak ada interaksi antara sisi pagar dan spesies pohon ( p = 0,127), sisi pagar dan TI ( p = 0,094) spesies pohon dan TI ( p = 0,975), atau di antara ketiga variabel bersama-sama ( p = 0,235), istilah interaksi ini kemudian dikeluarkan dari model akhir.
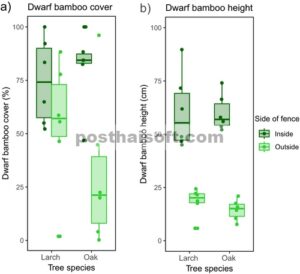
Rata-rata tegakan (a) tutupan dan (b) tinggi bambu kerdil yang diukur di 12 tegakan larch dan 12 tegakan oak di dalam dan di luar pagar rusa. Kotak-kotak menunjukkan rentang interkuartil (antara persentil ke-25 dan ke-75), dengan garis tengah menandai median (persentil ke-50). Kumis meluas ke titik data dalam 1,5 kali rentang interkuartil.
Dalam model kedua, kami malah bertujuan untuk menguji tinggi bambu kerdil sebagai variabel penjelas untuk kekayaan spesies bersama dengan variabel lainnya. Sama seperti pada model sebelumnya, tidak ada interaksi antara pasangan variabel: tinggi bambu kerdil dan spesies pohon ( p = 0,948), tinggi bambu kerdil dan TI ( p = 0,704), dan spesies pohon dan TI ( p = 0,924), atau di antara ketiga variabel secara bersamaan ( p = 0,216), jadi model dijalankan tanpa istilah interaksi ini. R2 marginal dan kondisional dihitung menggunakan fungsi r2_nakagawa() dari paket R “performance” (Lüdecke et al. 2021 ). Model akhir diperiksa untuk overdispersi.
Model-model yang berbeda dibandingkan menggunakan kriteria informasi Akaike (AIC; Lampiran S3 ). Selain model lengkap, dua model dengan nilai AIC terendah dipilih: satu menggunakan sisi pagar sebagai prediktor dan yang lainnya menggunakan tinggi bambu kerdil sebagai prediktor.
3.3 Analisis Multivariat
Komposisi komunitas spesies dianalisis dengan penskalaan multidimensi nonmetrik (NMDS) dalam paket R “vegan” (Oksanen et al. 2022 ), menggunakan indeks ketidakmiripan Bray–Curtis. Rata-rata tutupan spesies yang berbeda pada tingkat tegakan digunakan untuk analisis. Sebelum pengujian, spesies yang hanya muncul di satu plot dari seluruh survei dihapus dari kumpulan data. Secara total, 31 spesies dicatat hanya sekali, menyisakan 86 spesies untuk analisis.
Menggunakan solusi dua dimensi untuk NMDS menghasilkan nilai stres sebesar 0,196. Untuk memeriksa asosiasi tegakan dan spesies, kami membuat scatterplot dari dua sumbu NMDS pertama dari plot vegetasi, yang menunjukkan tegakan berpasangan untuk membandingkan kesamaan komunitas di seluruh pagar. Untuk menguji perbedaan komposisi komunitas di seluruh pagar dan antara spesies pohon tajuk, analisis varians multivariat permutasi (PERMANOVA) dilakukan menggunakan fungsi R adonis2() (Oksanen et al. 2022 ), menggunakan 999 permutasi. Karena tidak ada interaksi signifikan antara sisi pagar dan spesies pohon ( p = 0,495), model akhir dijalankan tanpa istilah interaksi.
Untuk menilai apakah pengecualian rusa atau kedekatan fisik antara plot berpasangan memiliki dampak yang lebih besar pada komposisi komunitas vegetasi lantai hutan, kami membandingkan jarak di antara skor situs NMDS. Sebagai ukuran dampak rusa, kami menggunakan jarak antara plot berpasangan di sisi pagar yang berlawanan, secara terpisah untuk plot dengan kanopi pohon ek dan larch. Untuk ukuran latar belakang variabilitas komunitas di antara plot pada sisi pagar tertentu, kami menghitung jarak rata-rata dari setiap plot ke semua plot dalam atau luar lainnya di ruang NMDS untuk plot kanopi pohon ek dan larch. Perbedaan jarak NMDS rata-rata di antara plot pagar dalam, melintang, dan luar yang dihitung diuji menggunakan uji- t sampel independen .
Terakhir, kami menggunakan analisis indikator spesies (analisis pola bertingkat) untuk memeriksa apakah beberapa spesies lebih terkait dengan satu sisi pagar. Analisis dilakukan menggunakan fungsi multipatt() dari paket R “Indicspecies” (De Caceres dan Legendre 2009 ). Spesies (tidak termasuk spesies tunggal) dianalisis untuk asosiasi kelompok (sisi pagar) dengan 9999 iterasi.
4 Hasil
4.1 Tutupan dan Tinggi Bambu Kerdil
Bambu kerdil memiliki tutupan yang jauh lebih tinggi di dalam pagar dibandingkan dengan di luar (Tabel 2 ). Di dalam pagar, tutupan rata-rata 77,8% ± 18,4% (semua nilai dilaporkan sebagai rata-rata ± SD) dibandingkan dengan 43,0% ± 33,6% di luar pagar (Gambar 4a ). Ketika menguji tutupan bambu kerdil secara terpisah untuk tegakan larch, tidak ada perbedaan yang signifikan antara bagian dalam (74,6% ± 20,1%) dan bagian luar (54,8% ± 30,1%) pagar (Tabel 2 ). Namun, pada tegakan ek, tutupan di dalam pagar (81,1% ± 17,9%) secara signifikan lebih besar daripada yang di luar (31,3% ± 35,5%; Tabel 2 ; Gambar 4a ).
TABEL 2. Perbedaan tutupan bambu kerdil, tinggi bambu kerdil, dan total tutupan tanaman vaskular di dalam dan luar pagar.
Variabel yang diuji Sisi pagar (dalam vs. luar): selisih rata-rata CI (95%) T df P
Penutupan bambu kerdil (%)
Semua stand 34.82 10,96–58,68 3.21 11 0,0083
Pohon Larch 19.85 -20,60 hingga 60,29 1.26 5 0.2628
Ek 49.79 15.34–84.24 3.72 5 0,0138
Tinggi bambu kerdil (cm)
Semua stand 43.91 34,72–53,09 10.52 11 < 0,0001
Pohon Larch 42.25 21,56–62,95 5.25 5 0,0033
Ek 45.57 37.16–53.97 13.94 5 < 0,0001
Total tutupan tumbuhan vaskular (%)
Semua stand 21.60 -10,72 hingga 53,92 1.47 11 0.1694
Pohon Larch -13,81 -45,99–18,35 -1,10 5 0.3198
Ek 57.01 13.20–100.82 3.35 5 0,0204 tahun
Catatan: Dalam analisis, rata-rata tingkat tegakan diuji menggunakan uji- t berpasangan .
Bambu kerdil juga secara signifikan lebih tinggi di dalam pagar (60,3 ± 15,0 cm) dibandingkan dengan di luar pagar (16,4 ± 6,7 cm; Tabel 2 ; Gambar 4b ). Perbedaan ini tetap konsisten saat menguji larch (60,6 ± 17,6 cm di dalam; 18,4 ± 6,6 cm di luar) dan tegakan ek secara terpisah (60,0 ± 8,5 cm di dalam; 14,4 ± 4,8 cm di luar; Tabel 2 ).
4.2 Kekayaan Spesies dan Tutupan Berbagai Kelompok Spesies
Sebanyak 117 spesies atau kelompok spesies tercatat di dalam plot: 11 pakis, 5 graminoid, 60 herba, 13 semak, 17 pohon, dan 11 tanaman merambat (Lampiran S1 : Tabel S1 ). Namun, graminoid (rumput dan alang-alang, termasuk bambu kerdil) tidak diidentifikasi hingga tingkat spesies, dan beberapa spesies pohon dan semak dicatat bersama-sama (lihat bagian Metode) sehingga jumlah total spesies diperkirakan secara konservatif. Jumlah total spesies tanaman adalah 82 di dalam pagar dan 92 di luar. Kekayaan spesies rata-rata 5,0 ± 3,2 spesies per plot dan 17,8 ± 8,4 spesies per tegakan di dalam pagar dan 10,4 ± 3,5 spesies per plot dan 28,6 ± 5,1 spesies per tegakan di luar pagar (Gambar 5a ).
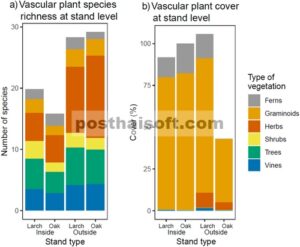
(a) Kekayaan spesies dan (b) tutupan berbagai kategori vegetasi lantai hutan di dalam dan luar pagar rusa pada 12 tegakan yang didominasi larch dan 12 tegakan yang didominasi oak. Estimasi kekayaan spesies bersifat konservatif karena graminoid dan beberapa spesies pohon dan semak hanya diidentifikasi pada tingkat genus.
Total tutupan tumbuhan vaskular tertinggi terdapat pada tegakan larch di luar pagar dan terendah pada tegakan oak di luar pagar (Gambar 5b ). Namun, total tutupan tumbuhan vaskular tidak berbeda secara signifikan ketika membandingkan semua tegakan di dalam (95,9% ± 31,9%) dan di luar (74,3% ± 50,0%) pagar (Tabel 2 ). Ketika tegakan yang didominasi larch pada sisi pagar yang berbeda dibandingkan, tidak ada perbedaan signifikan dalam total tutupan, tetapi untuk tegakan oak, total tutupan berbeda secara signifikan di seluruh pagar (Tabel 2 ).
Untuk kompilasi kekayaan spesies dan tutupan spesies berdasarkan tegakan, lihat Lampiran S2 dan Gambar S1a,b .
4.3 Dampak Pagar, Bambu Kerdil, dan Kepadatan Hutan terhadap Kekayaan Spesies
Analisis GLMM menunjukkan bahwa kekayaan spesies tingkat plot lebih tinggi di luar pagar daripada di dalam (Gambar 6 ; Tabel 3 : Model 1 dan 2). Ketika tinggi bambu kerdil digunakan sebagai prediktor dan bukan pagar, kekayaan spesies merespons secara negatif terhadap bambu kerdil yang lebih tinggi (Gambar 6 , Tabel 3 : Model 3 dan 4). Menggunakan tinggi bambu kerdil sebagai prediktor dan bukan sisi pagar juga memperbaiki model dengan menurunkan nilai AIC. Menambahkan pengaruh kerapatan hutan (seperti yang dihitung dengan indeks pengaruh pohon—TI) menghasilkan perbaikan model lebih lanjut, seperti yang ditunjukkan oleh AIC yang lebih rendah (lihat Lampiran S3 ). Nilai p yang signifikan untuk TI menunjukkan efek negatif pada kekayaan spesies ketika kerapatan hutan meningkat. Nilai AIC yang lebih rendah (1164,0 dan 1149,4) dalam Model 2 dan 4 menunjukkan bahwa memasukkan TI dan mengecualikan spesies pohon memberikan kecocokan model terbaik.
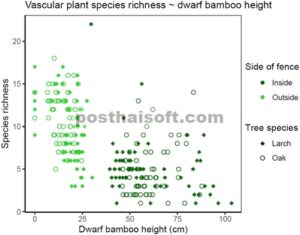
Kekayaan spesies tanaman vaskular di setiap petak sebagai fungsi dari tinggi bambu kerdil. Sepuluh petak didistribusikan di masing-masing dari 12 tegakan larch dan 12 tegakan oak di dalam dan di luar pagar rusa dengan total 240 petak.
TABEL 3. Hasil dari model GLMM (model lengkap dan model paling sesuai).
Model Kombinasi variabel penjelas Memperkirakan Bahasa Inggris dari P AIK Marjinal R 2 /kondisional R 2
1 (Mencegat) 1.546 0.117 Tanggal 13.22 < 0,0001 1166.0 0,411/0,632
Pagar (luar = 1) 0,775 tahun 0.109 Tanggal 7.11 < 0,0001
Spesies pohon (ek = 1) -0,004 0,144 tahun -0,03 0,976 tahun
Indeks pengaruh pohon -0,055 0,027 -2,04 0,041 tahun
2 (Mencegat) 1.544 0,093 16.63 < 0,0001 1164.0 0,411/0,632
Pagar (luar = 1) 0,775 tahun 0.109 Tanggal 7.11 < 0,0001
Indeks pengaruh pohon -0,055 0,027 -2,06 0,039
3 (Mencegat) 2.555 0.126 Tanggal 20.21 < 0,0001 1151.3 0,414/0,639
Tinggi bambu kerdil -0,016 0,002 -7,59 < 0,0001
Spesies pohon (ek = 1) -0,037 0.142 -0,26 0.794
Indeks pengaruh pohon -0,057 0,027 -2,08 0,037 hari
4 (Mencegat) 2.536 0.103 24.55 < 0,0001 1149.4 0,415/0,640
Tinggi bambu kerdil -0,016 0,002 -7,59 < 0,0001
Indeks pengaruh pohon -0,056 0,027 -2,07 0,039
Catatan: Dalam Model 1 (model lengkap), sisi pagar (kategori 1/0) digunakan sebagai prediktor kekayaan spesies bersama dengan spesies tajuk (larch/oak) dan indeks pengaruh pohon yang diskalakan ulang (berkisar dari -1,67 hingga 3,82). Dalam Model 2 (model paling sesuai saat sisi pagar digunakan sebagai prediktor), spesies pohon dihilangkan. Dalam Model 3, sisi pagar diganti dengan tinggi bambu kerdil (0–103,4 cm). Dalam Model 4 (model paling sesuai dengan tinggi bambu kerdil sebagai prediktor), spesies pohon dihilangkan. Tegakan dan pasangan tegakan (tegakan yang terletak di sisi berlawanan dari pagar) digunakan sebagai faktor efek acak dalam semua model.
4.4 Komposisi Komunitas Tumbuhan Vaskular
Komposisi komunitas tumbuhan vaskular dalam ruang NMDS secara signifikan berbeda di dalam dan di luar area berpagar (Tabel 4 ). Hal ini juga diilustrasikan dalam Gambar 7a , yang menunjukkan asosiasi spesies dengan tipe tegakan yang berbeda. Namun, tidak ada perbedaan NMDS yang signifikan antara komunitas di bawah pohon larch dan pohon oak (Tabel 4 ; Gambar 7a ). Gambar 7b , yang didasarkan pada perhitungan yang sama, malah menunjukkan asosiasi antara tegakan yang terletak di sisi berlawanan dari pagar. Untuk vegetasi tingkat bawah di bawah tajuk pohon oak, jarak ruang NMDS melintasi pagar antara pasangan plot secara signifikan lebih besar daripada jarak NMDS rata-rata di antara plot baik di dalam maupun di luar pagar (Gambar 8 ; uji- t , p < 0,05). Untuk plot di bawah tajuk pohon larch, jarak NMDS antar pagar secara signifikan lebih kecil dibandingkan dengan jarak NMDS antar plot di luar pagar (Gambar 8 ; uji- t , p < 0,05), namun tidak dapat dibedakan dari jarak NMDS antar plot di luar pagar (Gambar 8 ; uji- t , p > 0,05).
TABEL 4. Analisis varians multivariat permutasi (PERMANOVA) dari matriks spesies tumbuhan vaskular.
df Jumlah kuadrat R 2 F P
Spesies pohon 1 0,179 tahun 0,051 tahun 1.371 0.219
Sisi pagar 1 0,598 0.170 4.589 0,001
Sisa 21 2.738 0,779 tahun
Total 23 3.515 1.000
Catatan: Pengujian dijalankan dengan 999 iterasi.
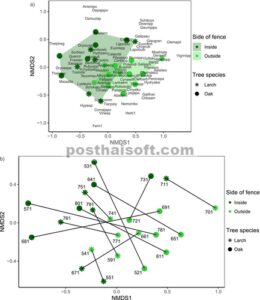
Biplot ordinasi NMDS menunjukkan komposisi komunitas tumbuhan vaskular di dua belas tegakan larch dan dua belas tegakan oak. Tegakan tersebut terletak di dalam (hijau tua) dan di luar (hijau muda) pagar rusa. Plot (a) menunjukkan asosiasi spesies tumbuhan bawah dengan berbagai jenis tegakan. Plot (b) menunjukkan kesamaan antara tegakan berpasangan, di mana garis menghubungkan tegakan berpasangan di sisi pagar yang berlawanan. Angka-angka dalam plot adalah nomor tegakan yang digunakan dalam peta pada Gambar 1 .
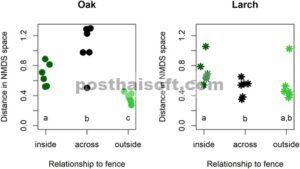
Kesamaan vegetasi di petak-petak di bawah tajuk pohon ek dan larch dan bagaimana hal itu dipengaruhi oleh pengecualian rusa. Jarak NMDS yang lebih rendah menunjukkan kesamaan yang lebih besar. Untuk kategori dalam dan luar, titik-titik menunjukkan jarak NMDS rata-rata dari setiap petak ke semua petak lain dengan jenis pohon tajuk yang sama dan sisi pagar. Untuk kategori melintang, setiap nilai berhubungan dengan sepasang petak dengan komposisi tajuk yang sama, yang secara fisik terletak berdekatan satu sama lain tetapi pada sisi pagar yang berlawanan. Huruf yang berbeda menunjukkan uji- t yang signifikan secara statistik pada p < 0,05.
Analisis spesies indikator menunjukkan bahwa secara total, 13 spesies atau kelompok spesies menunjukkan hubungan yang signifikan ( p < 0,05) dengan satu sisi pagar (Tabel 5 ). Satu spesies secara signifikan lebih umum di dalam pagar, dan 12 secara signifikan lebih umum di luar pagar.
TABEL 5. Spesies atau kelompok spesies yang menunjukkan hubungan signifikan dengan kedua sisi pagar dan nilai indikatornya yang mewakili kekuatan hubungan tersebut.
Nilai indikator P
Di dalam pagar
Jamur Dryopteris crassirhizoma 0.645 0,0385
Pagar luar
Aster leiophyllus var. leiofilus 0,957 tahun 0,0001
Athyrium spp. 0,735 0,0156
Carex spp. 0.943 0,0079 pukul 0,0079
Chamaele decumbens var. decumbens 0.697 0,0373 pukul 0,0373
Clinopodium micranthum var. micranthum / C. multicaule var. yakusimense 0.707 0,0132
Cynanchum caudatum / C.wilfordii 0.764 0,0040
Kalopanax septemlobus subsp. septemlobus 0.870 0,0012
Persikaria debilis 0.726 0,0174 tahun
Poaceae spp. 0.762 0,0221
Potentilla indica 0.804 0,0049 pukul 0,0049
Thalictrum spp. 0.748 0,0237
Viola spp. 0.921 0,0005
5 Diskusi
Kelimpahan rusa saat ini di sebagian besar belahan bumi utara telah menyebabkan tekanan penggembalaan yang tinggi yang sangat memengaruhi flora hutan di beberapa area. Penelitian ini bertujuan untuk meneliti dampak pada flora setelah 21 tahun rusa sika dilarang masuk di Taman Nasional Nikko. Pagar larangan masuk rusa mengakibatkan peningkatan signifikan pada tinggi bambu kerdil di dalam pagar dan tutupan yang tinggi, yang menunjukkan pemulihannya di area yang dilindungi. Namun, tutupan bambu kerdil menunjukkan variasi di antara tegakan di luar pagar, yang mungkin dipengaruhi oleh preferensi rusa, topografi lokal, dan durasi tutupan salju.
Studi kami menunjukkan bahwa dominasi bambu kerdil di dalam pagar menyebabkan berkurangnya kekayaan spesies tumbuhan berpembuluh. Temuan ini untuk pertama kalinya mempertimbangkan efek ini di berbagai jenis hutan dan kerapatan hutan. Kami tidak menemukan perbedaan dalam kekayaan spesies dan komposisi komunitas tumbuhan berpembuluh antara tegakan yang didominasi larch dan oak. Meskipun kami mengamati efek lemah kerapatan tegakan pada kekayaan spesies, faktor utama yang memengaruhi komunitas spesies lantai hutan tampaknya adalah pengecualian rusa di dalam pagar dan penggembalaan intensif di luar pagar. Pengecualian rusa juga mengakibatkan perkembangan komposisi komunitas yang berbeda di dalam pagar dibandingkan dengan vegetasi yang terpengaruh oleh penggembalaan di luar pagar. Di bawah ini, kami membahas hasil ini dalam konteks konservasi flora hutan dengan memagari di dalam ekosistem hutan yang terpengaruh oleh penggembalaan rusa yang intensif.
5.1 Bambu Kerdil
Temuan kami menunjukkan efek yang jelas dari pengecualian rusa pada kelimpahan bambu kerdil. Hasil ini sejalan dengan penelitian sebelumnya, yang menyoroti pemulihan cepat oleh bambu kerdil di daerah yang sebelumnya terganggu dan kemampuannya untuk bangkit kembali ketika dilindungi dari penggembalaan rusa yang berat (Itô dan Hino 2007 ; Shibata et al. 2008 ; Kisanuki et al. 2009 ; Kudo et al. 2017 ). Dalam penelitian kami, perbedaan antara sisi pagar paling menonjol dalam hal tinggi bambu kerdil. Bambu kerdil yang lebih tinggi yang ditemukan di dalam pagar menunjukkan bahwa pengecualian rusa dengan pagar telah memungkinkan pertumbuhan bambu kerdil yang lebih besar. Rusa yang merumput di luar pagar, sebaliknya, telah menghasilkan bambu kerdil yang lebih pendek.
Ada juga variasi besar dalam tutupan tanah di antara tegakan yang berbeda di luar pagar (Lampiran S2 , Gambar S1b ). Khususnya, tutupan bambu kerdil serupa antara tegakan larch di sisi pagar yang berbeda tetapi secara signifikan lebih tinggi di dalam pagar jika dibandingkan di antara semua tegakan bersama-sama dan di tegakan ek secara terpisah. Tutupan bambu kerdil khususnya rendah di satu tegakan larch dan di satu tegakan ek di luar pagar. Di tegakan larch, lapisan bambu kerdil tampaknya digantikan oleh Carex dan Aster leiophyllus var. leiophyllus . Perubahan dominasi ini mungkin disebabkan oleh penggembalaan rusa sika dan kemungkinan gangguan lainnya, yang memengaruhi prevalensi bambu kerdil. Tampaknya A. leiophyllus var. leiophyllus , yang tidak dapat dimakan rusa, tumbuh subur di tegakan di luar pagar; oleh karena itu, kemungkinan besar disukai oleh tekanan penggembalaan rusa yang tinggi di area tersebut (Nomiya et al. 2003 ; Tamura 2019 ). Lebih jauh lagi, tidak hanya tutupan bambu kerdil tetapi juga total tutupan tanaman vaskular sangat rendah di satu tegakan pohon ek di area curam di luar pagar. Di sini, kombinasi lereng curam dan penggembalaan rusa mungkin telah memengaruhi stabilitas tanah (Ohashi et al. 2014 ), sehingga berdampak buruk pada vegetasi lantai hutan.
Variasi dalam tutupan bambu kerdil dan total tutupan tanaman vaskular antara lokasi di luar pagar juga dapat dipengaruhi oleh topografi lokal, misalnya, oleh pengaruh pada naungan, kadar air tanah, dan umur tutupan salju. Pemilihan habitat rusa, yang dipengaruhi oleh ketersediaan dan kualitas hijauan serta tutupan untuk keamanan, juga dapat memengaruhi pola penggembalaan temporal dan spasial (Godvik et al. 2009 ). Ini dapat membantu menjelaskan perbedaan yang jelas dalam tutupan bambu kerdil yang diamati di antara tegakan di luar pagar. Khususnya, mengingat perbedaan yang konsisten dalam tinggi bambu kerdil dibandingkan dengan hasil tutupan yang tidak konsisten, tinggi bambu kerdil dapat berfungsi sebagai indikator tekanan penggembalaan rusa yang lebih andal daripada tutupan saja.
5.2 Kekayaan Spesies Tumbuhan Vaskular Terkait dengan Bambu Kerdil
Analisis mengungkapkan bahwa kekayaan spesies tanaman vaskular secara konsisten lebih tinggi di luar pagar dibandingkan dengan di dalam, terlepas dari apakah tegakan tersebut didominasi oleh larch atau oak (Gambar 6 ; Tabel 3 ). Lebih jauh lagi, hubungan negatif diamati antara kekayaan spesies dan peningkatan tinggi bambu kerdil, dan tinggi bambu kerdil adalah prediktor yang lebih baik untuk kekayaan spesies daripada hanya sisi pagar (Gambar 6 ; Tabel 3 ). Dominasi bambu kerdil, yang mengarah pada pengurangan kekayaan tanaman vaskular, sebelumnya telah didokumentasikan di berbagai daerah di Jepang (Fukamachi et al. 1996 ; Konno 2002 ; Itô dan Hino 2005 ). Mekanisme yang terlibat dalam pengecualian tanaman lain ini sering kali memerlukan naungan, jatuhnya serasah, dan berkurangnya kadar air tanah (Doležal et al. 2009 ). Respons terhadap kemampuan kompetitif ini mungkin berbeda di antara spesies tanaman tingkat bawah. Secara khusus, tinggi batang tampaknya menjadi prediktor penting kekayaan spesies herba, karena bambu kerdil yang lebih tinggi lebih cenderung menaungi vegetasi lain (Kudo et al. 2011 ). Di lokasi penelitian ini, tanaman bambu kerdil dengan tinggi antara 25 dan 45 cm cukup jarang (perhatikan celah pada Gambar 6 ), jadi tidak memungkinkan untuk mengambil sampel komunitas spesies lantai hutan yang berbeda yang tumbuh di antara bambu kerdil dengan ketinggian ini.
Selain penurunan kekayaan spesies yang diamati di dalam pagar akibat bambu kerdil yang lebih tinggi, model kekayaan spesies juga mengungkap penurunan kekayaan spesies yang terkait dengan kepadatan tegakan yang lebih tinggi, meskipun dampaknya hanya sedikit signifikan dalam model. Khususnya, tidak ada dampak spesies pohon pada kekayaan spesies. Mungkin, dampak spesies pohon dan kepadatan pohon di sini lebih kecil daripada dampak penggembalaan intensif di luar pagar dan pengecualian bambu kerdil kompetitif di dalam pagar, yang tampaknya menjadi pengaruh utama pada komunitas lantai hutan.
5.3 Komposisi Komunitas
Analisis komposisi komunitas mengungkap perbedaan yang nyata antara tegakan di dalam dan di luar pagar (Gambar 7a ; Tabel 4 ). Menariknya, tidak ada perbedaan signifikan dalam lokasi komposisi komunitas di ruang NMDS antara tegakan yang didominasi larch dan oak. Jarak NMDS antara tegakan larch yang berpasangan di sisi berlawanan dari pagar tidak berbeda secara signifikan dari variasi antara tegakan di luar pagar (Gambar 8 ). Ini menunjukkan bahwa komunitas di tegakan larch di seberang pagar lebih mirip dibandingkan dengan variasi di antara plot larch di luar pagar. Untuk tegakan oak yang berpasangan, variasi berbeda secara signifikan antara pasangan dan tegakan di luar dan di dalam pagar. Ini menunjukkan bahwa ada variasi yang lebih besar antara komunitas tanaman di seberang pagar daripada ketika membandingkan komunitas di luar atau di dalam pagar satu sama lain.
Sebanyak 13 spesies diasosiasikan dengan bagian luar pagar. Di antara mereka, herba Aster leiophyllus var. leiophyllus dan Viola spp., bersama dengan tanaman merambat Actinidia arguta , Cynanchum caudatum , dan C. wilfordii , sangat melimpah. Dua spesies pohon yang melimpah diasosiasikan dengan bagian luar pagar adalah Acer rufinerve dan Kalopanax septemlobus subsp. septemlobus . Karena A. rufinerve tidak enak dimakan (Sakata et al. 2021 ), ini adalah satu-satunya spesies pohon yang tampaknya memiliki potensi untuk membentuk lapisan pohon baru di masa mendatang. K. septemlobus subsp. septemlobus hanya ditemukan sebagai bibit kecil. Karena spesies ini disukai oleh rusa, spesies ini tidak mungkin tumbuh besar tanpa perlindungan dari pemangsaan (Takahashi dan Kaji 2001 ). Di kedua sisi pagar, terdapat banyak spesies pohon termasuk Acer spp., Malus toringo , dan Quercus crispula . Namun, mereka hanya hadir dalam bentuk bibit kecil, yang sering kali tersembunyi di bawah bambu kerdil yang tinggi di dalam area berpagar.
Spesies pohon lain yang tumbuh dalam jumlah yang cukup banyak di kedua sisi pagar adalah berbagai spesies Acer spp., Malus toringo , dan Quercus crispula . Kesamaan dari spesies-spesies ini adalah bahwa mereka semua tumbuh sebagai bibit kecil, baik tersembunyi di bawah bambu kerdil yang tinggi di dalam pagar atau belum ditemukan oleh rusa sika di luar pagar. Selain itu, pohon ek muda dapat ditemukan dalam bentuk pengisap kerah akar yang tumbuh tepat di samping pohon induk. Meskipun demikian, selain Acer rufinerve yang tidak dapat dimakan , kami melihat kurangnya regenerasi pohon yang berhasil di kedua sisi pagar. Tidak adanya perekrutan bibit pohon baru dari spesies yang saat ini hanya tumbuh sebagai pohon besar dapat menimbulkan tantangan jangka panjang untuk melestarikan hutan di taman nasional.
Kurangnya regenerasi pohon tidak semata-mata disebabkan oleh tekanan penjelajahan yang tinggi di luar pagar. Menariknya, kegagalan regenerasi tampaknya sama parahnya di dalam area berpagar, di mana bibit muda berjuang untuk bertahan hidup karena pertumbuhan yang kompetitif dan dominasi lapisan bambu kerdil yang tinggi dan menaungi. Pengamatan ini sejalan dengan penelitian sebelumnya yang telah membahas efek pagar pada regenerasi pohon (Itô dan Hino 2007 ; Doležal et al. 2009 ; Murata et al. 2009 ; Simončič et al. 2019 ). Sebaliknya, Akashi et al. ( 2021 ) menemukan bahwa bambu kerdil yang tinggi dapat melindungi bibit pohon dengan menyembunyikannya dari rusa. Efek persembunyian bambu kerdil adalah sesuatu yang tidak dapat kami evaluasi dalam penelitian ini, karena bambu kerdil yang tinggi hanya ada di area yang tidak didatangi rusa. Lebih banyak upaya untuk mempromosikan regenerasi pohon guna mengamankan pengembangan hutan berlapis-lapis alami, alih-alih hutan yang kurang lebih satu lapis saat ini, diperlukan di area ini. Salah satu metode potensial untuk meningkatkan keanekaragaman flora hutan dan memfasilitasi regenerasi pohon adalah penebangan bambu kerdil secara eksperimental di area tertentu. Metode ini sebelumnya telah berhasil diuji untuk memulihkan flora hutan pegunungan (Kudo et al. 2017 ).
Hanya satu spesies yang signifikan sebagai indikator untuk bagian dalam kandang rusa: spesies pakis besar berbentuk kerucut Dryopteris crassirhizoma . Spesies ini memiliki daun yang mampu melewati musim dingin, yang membuatnya sangat diminati oleh rusa sebagai makanan musim dingin. Namun, spesies ini telah menunjukkan ketahanan terhadap penggergajian menurut Inatomi et al. ( 2022 ). Khususnya, daun yang mampu melewati musim dingin tidak hanya memberi pakis awal yang baik di musim semi tetapi juga memberikan naungan yang penting, memungkinkannya untuk mendominasi area di sekitarnya, memfasilitasi perkembangan daun baru (Tani dan Kudo 2005 ). Strategi ini mungkin sangat berhasil untuk menahan dominasi bambu kerdil, berkontribusi pada keberhasilan D. crassirhizoma bahkan di antara bambu kerdil yang tinggi.
5.4 Peringatan dan Potensi untuk Studi Masa Depan
Dalam penelitian ini, kami menyelidiki dua habitat yang kontras dalam hal penggembalaan rusa: satu mengalami tekanan penggembalaan yang sangat tinggi dan yang lainnya di mana rusa dikecualikan. Temuan kami menunjukkan bahwa kedua ekstrem tersebut menghadirkan tantangan dalam mendukung komunitas tumbuhan vaskular lantai hutan yang beragam. Namun, aspek penting yang masih belum dieksplorasi dalam penelitian ini adalah dampak tingkat penggembalaan rusa menengah terhadap vegetasi lantai hutan. Ini berarti kami tidak dapat menarik kesimpulan tentang tingkat tekanan penggembalaan mana yang optimal untuk mendukung flora lantai hutan yang kaya.
Keterbatasan tambahan dari studi ini adalah tidak adanya data historis mengenai vegetasi lantai hutan sebelum peningkatan signifikan populasi rusa selama tahun 1980-an. Pemantauan jangka panjang terhadap flora di area ini, serta di wilayah lain dengan populasi rusa yang besar, sangat penting untuk memahami dampak kepadatan rusa dari waktu ke waktu. Menyertakan data historis mengenai flora hutan akan memungkinkan evaluasi yang lebih langsung terhadap perubahan bunga selama bertahun-tahun. Mengatasi peringatan ini dan melakukan studi di masa mendatang yang mencakup tekanan penggembalaan rusa yang lebih luas dan menggabungkan data historis akan berkontribusi pada pemahaman yang lebih komprehensif tentang dinamika rumit antara dampak perburuan rusa dan keanekaragaman vegetasi lantai hutan.
Saat mengamati vegetasi di bawah tajuk, kami memusatkan plot di sekitar pepohonan. Pendekatan ini mungkin menghasilkan hasil yang sedikit berbeda dibandingkan dengan mendistribusikan plot di ruang antara pepohonan, karena peningkatan naungan langsung di bawah tajuk mungkin memengaruhi vegetasi. Namun, saat membandingkan flora yang lebih jauh dari pepohonan, kami tidak mengamati perbedaan yang nyata dalam kekayaan spesies atau komposisi komunitas.
5.5 Implikasi bagi Konservasi Alam
Karena penelitian kami dilakukan 21 tahun setelah pagar dipasang, suksesi mungkin masih berlangsung, dengan vegetasi di dalam pagar masih pulih dari perburuan rusa yang intens di masa lalu. Kami dapat melihat dengan jelas bahwa bambu kerdil adalah pemenangnya pada tahap suksesi ini. Namun, perspektif ekologi jangka panjang mungkin adalah bahwa dominasi bambu kerdil atas spesies lantai hutan lainnya dapat berubah. Pembungaan massal bambu kerdil biasanya terjadi setiap 60–100 tahun atau terkadang lebih lama (Makita 1998 ). Pembungaan dan penyemaian massal diikuti oleh mati pucuk yang luas. Penuaan monokarpik dapat ditemukan di berbagai jenis ekosistem hutan (Tsvuura et al. 2011 ) dan merupakan gangguan penting di lantai hutan yang didominasi bambu kerdil, yang juga membantu dalam membentuk struktur hutan karena penuaan bambu kerdil yang sinkron biasanya diikuti oleh perkecambahan biji dari bank biji (Taylor et al. 1995 ). Pemicu pembungaan massal dan lamanya interval antara pembungaan masih belum dipahami (Cho et al. 2017 ). Dengan mempertimbangkan perspektif yang lebih panjang ini, flora saat ini yang sangat tertekan oleh dominasi bambu kerdil dapat berubah dengan cepat saat bambu kerdil berbunga. Namun, karena pembungaan massal dan mati pucuk bambu kerdil sangat jarang terjadi, upaya konservasi mungkin juga diperlukan untuk memastikan bahwa spesies hutan, terutama tanaman semusim dan spesies dengan persediaan benih terbatas, dapat bertahan hidup dan bereproduksi dalam jangka panjang. Penelitian lain telah menunjukkan bahwa penebangan bambu kerdil untuk mengurangi persaingan dapat berhasil meningkatkan keanekaragaman tumbuhan berpembuluh (Kudo et al. 2017 ). Lebih banyak penelitian tentang penebangan bambu kerdil untuk tujuan konservasi flora direkomendasikan.
Masalah penggembalaan rusa sika yang luas bukan hanya fenomena lokal di dalam dan sekitar Nikko; kerusakan rusa sika terhadap flora dan regenerasi hutan telah dilaporkan di seluruh Jepang, dan spesies rusa lainnya menyebabkan masalah bagi konservasi alam di banyak wilayah di belahan bumi utara. Upaya pengelolaan rusa saat ini jelas tidak cukup untuk mempertahankan keanekaragaman flora dan memungkinkan regenerasi hutan alami. Populasi rusa yang lebih kecil diperlukan sebelum keanekaragaman herba hutan habis, yang menyebabkan efek yang tidak dapat dipulihkan, termasuk hilangnya spesies tanaman vaskular lokal dengan adaptasi lokal. Diperlukan pengelolaan permainan yang lebih baik dan penentuan prioritas area untuk upaya konservasi yang ditargetkan.
Kontribusi Penulis
Lisa Petersson, Carl Salk, Daniel Jensen, Göran Thor, dan Tatsuhiro Ohkubo merancang penelitian dan menulis artikel. Lisa Petersson dan Daniel Jensen melakukan inventarisasi dan pengukuran lapangan. Lisa Petersson, Carl Salk, dan Daniel Jensen menganalisis data dan menyusun angka.
Ucapan Terima Kasih
Kami berterima kasih kepada Kantor Taman Nasional Nikko yang telah menyediakan peta dan izin untuk melakukan penelitian ini. Kami juga berterima kasih kepada Kantor Distrik Hutan Badan Kehutanan di Nikko yang telah mengizinkan masuk ke area penelitian dan bantuan mereka. Kami sangat berterima kasih kepada Bapak Yoshikawa Makoto atas keahliannya yang tak ternilai dalam mengidentifikasi spesies tanaman. Kami juga ingin berterima kasih kepada Dr. Aizawa Mineaki di Laboratorium Ekologi Hutan dan Silvikultur di Departemen Ilmu Kehutanan, Universitas Utsunomiya serta Dr. Ishiguri Futoshi dan Dr. Nezu Ikumi di Laboratorium Ilmu Material Kayu di Departemen Ilmu Kehutanan, Universitas Utsunomiya atas penggunaan fasilitas laboratorium. Terima kasih juga kepada Bapak Sawada Tadashi di Nikko Shizen Freai House, Universitas Utsunomiya, yang telah menyediakan akomodasi selama kerja lapangan. Lisa Petersson didanai oleh beasiswa pascadoktoral Japan Society for the Promotion of Science (JSPS) (PE20742). Tinggalnya Göran Thor di Jepang pada tahun 2022 diberikan oleh beasiswa BRIDGE (BR220502) dari Masyarakat Jepang untuk Pengembangan Ilmu Pengetahuan (JSPS).
Konflik Kepentingan
Penulis menyatakan tidak ada konflik kepentingan.