
Abstrak
idTautan ini menuju ke bagian bahasa InggristinggiTautan ini menuju ke bagian Latvia
Hubungan lintas kerajaan memainkan peran mendasar dalam proses ekologi. Namun, pemahaman kita tentang keberadaan jamur dan tumbuhan di hutan hujan tropis dan dampak potensial perubahan penggunaan lahan yang membentuk hubungan spesies masih terbatas.
Dengan menggunakan pengurutan amplikon pada DNA dari akar dan komunitas jamur terkaitnya, kami bertujuan untuk memahami dampak transformasi hutan hujan terhadap komposisi dan struktur jaringan ekologi akar–jamur di lanskap yang dimodifikasi manusia di Sumatra, Indonesia.
Setiap jenis penggunaan lahan mendukung serangkaian spesies indikator yang khas, yaitu organisme yang mencerminkan kondisi lingkungan tertentu dan dapat menandakan perubahan dalam kesehatan ekosistem. Kami mengamati penurunan kekayaan taksa tanaman indikator dan asosiasi tanaman-jamur seiring dengan meningkatnya intensifikasi penggunaan lahan. Selain itu, terjadi pergantian komunitas akar, bergeser dari spesies asli dan endemik di hutan hujan ke spesies herba generalis non-asli di perkebunan karet dan kelapa sawit.
Konektivitas tumbuhan-jamur menurun secara signifikan seiring dengan meningkatnya intensifikasi penggunaan lahan, yang menunjukkan bahwa ekosistem yang dikelola mungkin telah melemahkan interaksi akar-jamur. Analisis jaringan menyoroti respons yang berbeda dari berbagai kelompok jamur. Misalnya, jamur mikoriza arbuskular (AMF) menunjukkan lebih sedikit koneksi dengan modul yang terkait dengan akar kelapa sawit dan karet, yang menunjukkan melemahnya asosiasi akar-jamur dalam monokultur. Hal ini sejalan dengan pengurangan yang diamati dalam keanekaragaman AMF di area penggunaan lahan yang dikonversi dibandingkan dengan hutan, yang selanjutnya memperkuat dampak negatif dari praktik penggunaan lahan dalam monokultur kelapa sawit dan karet terhadap keanekaragaman AMF.
Sintesis . Pengukuran dampak transformasi hutan hujan di bawah tanah dibatasi oleh pemahaman kita tentang kelompok fungsional jamur. Sistem yang sangat dimodifikasi menunjukkan lebih sedikit koneksi, yang menunjukkan restrukturisasi dinamis hubungan akar-jamur sebagai respons terhadap perubahan penggunaan lahan. Memahami interaksi rumit antara tanaman dan jamur dalam menghadapi perubahan penggunaan lahan dapat memberikan informasi berharga untuk upaya konservasi, praktik pertanian, dan strategi pengelolaan ekosistem yang bertujuan untuk meningkatkan keanekaragaman hayati, kesehatan tanah, dan ketahanan ekosistem dalam konteks perubahan kondisi lingkungan. Selain itu, hal ini menggarisbawahi pentingnya jaringan masyarakat dalam perencanaan penggunaan lahan dan keputusan pengelolaan untuk mendukung keanekaragaman tanaman dan jamur dalam ekosistem terestrial.
1. PENDAHULUAN
Hutan hujan tropis adalah ekosistem yang sangat beragam yang mengandung beragam habitat mikro dan organisme (Eiserhardt et al., 2017 ; Wardle et al., 2004 ). Namun, hutan hujan menghadapi ancaman besar karena penggantiannya yang cepat dengan perkebunan tanaman komersial seperti kelapa sawit dan karet (Rembold et al., 2017 ; Zemp et al., 2023 ). Misalnya, perluasan pertanian yang sedang berlangsung telah menghasilkan sekitar 21 juta hektar perkebunan kelapa sawit secara global (Descals et al., 2021 ; Tedersoo et al., 2014 ). Secara keseluruhan, keragaman taksonomi spesies asli telah menurun akibat perubahan penggunaan lahan (Ballauff et al., 2021 ; Barnes et al., 2017 ; Brinkmann et al., 2019 ; Felipe-Lucia et al., 2020 ; Newbold et al., 2016 ). Namun, untuk lebih memahami berbagai dampak perluasan pertanian pada ekosistem tropis, kita sangat perlu memperkuat pengetahuan kita tentang dampak alih fungsi lahan pada interaksi spesies (Brinkmann et al., 2019 ; Felipe-Lucia et al., 2020 ; Newbold et al., 2016 ; Romdhane et al., 2022 ).
Asosiasi tumbuhan–jamur memainkan peran mendasar dalam membentuk struktur dan fungsi ekosistem tropis (Põlme et al., 2018 ; Tedersoo et al., 2014 , 2022 ), memberikan wawasan tentang ketahanan, kapasitas adaptif, dan kesehatan ekosistem hutan (Trivedi et al., 2020 ). Interaksi akar–mikroba yang melibatkan berbagai kingdom dan serikat fungsional (Brunel et al., 2020 ; Ferlian et al., 2018 ; McLaren & Callahan, 2020 ; Trivedi et al., 2020 ; Vorburger & Perlman, 2018 ) telah dikaitkan dengan preferensi mutualistik, perbedaan evolusi dan sifat di antara inang, serta keterkaitan filogenetik dan pengecualian kompetitif antara jamur (Alzarhani et al., 2019 ; Francioli et al., 2021 ). Misalnya, kelimpahan dan keanekaragaman jamur mikoriza arbuskular (AMF) telah ditemukan merespons perubahan dalam keanekaragaman dan komposisi tanaman (Deyn et al., 2011 ; Gui et al., 2017 ), dengan preferensi inang–AMF kemungkinan dimodulasi oleh gugus fungsi tanaman, alih-alih mencerminkan interaksi spesies individu (Edy et al., 2022 ; Francioli et al., 2021 ; Zanne et al., 2020 ). Selain itu, peningkatan kelimpahan AMF di rizosfer diperkirakan akan mendorong penurunan kelimpahan patogen (Francioli et al., 2021 ; Sweeney et al., 2021 ; Zanne et al., 2020 ) dan berkorelasi positif dengan kelimpahan jamur saprotrofik (Francioli et al., 2021 ; van der Heijden & Hartmann, 2016 ). Meskipun di hutan dataran rendah tropis, sebagian besar spesies pohon berasosiasi dengan AMF, spesies tanaman dalam famili Dipterocarpaceae dan Fagaceae diketahui mengandung jamur ektomikoriza (EMF; Bahram et al., 2014 ; Matchado et al., 2021 ). Meskipun demikian, pemahaman kita tentang asosiasi akar-jamur di hutan hujan tropis dataran rendah sebagian besar berasal dari studi yang berkonsentrasi pada akar-AMF (Gui et al., 2017 ; Koorem et al., 2017 ), dengan studi EMF terutama dari ekosistem beriklim sedang (Encinas-Viso et al., 2016 ; Hogan et al., 2023 ; Kuang et al., 2021 ; Van Geel et al., 2018 ).
Konversi hutan hujan tropis dalam skala besar menjadi monokultur mengurangi keanekaragaman mikrohabitat, yang menyebabkan pergantian spesies (Ballauff et al., 2021 ; Barnes et al., 2017 ; Brinkmann et al., 2019 ) dan hilangnya keanekaragaman tumbuhan dan jamur (Barnes et al., 2017 ). Misalnya, ketika menganalisis asosiasi akar–AMF spesifik spesies dalam sampel akar individual, terdapat penurunan signifikan kekayaan AMF pada akar kelapa sawit dan karet dibandingkan dengan akar dari tanaman yang diambil sampelnya di hutan hujan (Edy et al., 2022 ). Filter lingkungan yang terkait dengan konversi hutan dapat memilih spesies yang toleran dan oportunistik (tanaman dan jamur) yang dapat menempati rentang relung yang besar dan mengalahkan spesies asli di lanskap yang dikonversi (Gioria et al., 2023 ; Hulme, 2017 ; McLaren & Callahan, 2020 ; Nayanakantha, 2007 ; Vilà & Weiner, 2004 ; Walther et al., 2009 ), yang mengakibatkan perubahan dalam komposisi komunitas. Oleh karena itu, wawasan untuk memahami interaksi tingkat komunitas dalam ekosistem alami dan yang dimodifikasi manusia sangat dibutuhkan (Chen et al., 2021 ; Delavaux et al., 2017 ; Sepp et al., 2019 ), namun studi dalam hotspot keanekaragaman tropis pada asosiasi di bawah tanah masih jarang.
Dengan menggabungkan pengurutan amplikon pada DNA dari sampel identik untuk penilaian komunitas tumbuhan dan jamur dengan analisis jaringan, kami dapat memberikan wawasan berharga ke dalam mekanisme yang mendorong koeksistensi spesies dari berbagai kerajaan dalam komunitas biologis, mengungkap interaksi trofik dan non-trofik dalam kondisi lingkungan tertentu (Grass et al., 2020 ; Montoya-Sánchez et al., 2023 ; Tiede et al., 2016 ; Zemp et al., 2023 ). Analisis jaringan memainkan peran penting dalam mengidentifikasi node yang sangat terhubung, memfasilitasi identifikasi spesies kunci dan interaksi yang penting untuk membentuk struktur komunitas (Guimerà & Amaral, 2005 ; Põlme et al., 2018 ; van der Heijden & Hartmann, 2016 ). Pendekatan yang efektif untuk mengidentifikasi spesies khusus penggunaan lahan (yaitu spesies indikator) adalah penggunaan indeks nilai indikator (IndVal), yang mengevaluasi pola distribusi yang paling sesuai dengan taksa yang dimaksud (De Cáceres et al., 2010 ), menentukan apakah suatu spesies khusus untuk penggunaan lahan tertentu atau terkait dengan beberapa jenis penggunaan lahan.
Di sini, dengan menggunakan kumpulan data yang belum pernah ada sebelumnya untuk daerah tropis yang berisi komunitas akar dan jamur dari empat jenis penggunaan lahan di Sumatra, Indonesia, kami bertujuan untuk memahami dampak transformasi hutan hujan pada komposisi dan struktur jaringan akar-jamur di hotspot keanekaragaman hayati tropis global. Untuk melakukannya, pertama-tama kami mengidentifikasi pola dalam komunitas akar-jamur dan memperkirakan spesies indikator yang terkait dengan setiap jenis penggunaan lahan. Kedua, kami memperkirakan asosiasi akar-jamur melalui analisis jaringan ko-kemunculan. Ketiga, kami memeriksa taksa yang sangat terhubung yang membentuk komposisi dan struktur jaringan ekologi. Kami berhipotesis bahwa transformasi hutan merupakan pendorong utama asosiasi akar-jamur melalui perubahan dalam keanekaragaman, komposisi, dan sifat akar (Ballauff et al., 2021 ). Perkebunan, misalnya, memiliki 59% spesies tanaman lebih sedikit daripada hutan hujan, sedangkan yang terakhir sebagian besar terdiri dari spesies asli, termasuk spesialis hutan, versus kekayaan dan kelimpahan spesies non-asli yang lebih tinggi di jenis penggunaan lahan lainnya (Rembold et al., 2017 ). Dengan demikian, kami menduga plot hutan hujan dikaitkan dengan akar dari spesies langka dan endemik. Sebaliknya, akar dari spesies non-asli dan generalis, bersama dengan kelompok jamur yang beradaptasi untuk bertahan atau tumbuh subur di relung yang sangat terganggu, dapat berfungsi sebagai indikator intensitas penggunaan lahan karena ketahanannya terhadap kondisi yang merugikan. Akhirnya, di seluruh jenis penggunaan lahan, kami berhipotesis bahwa taksa yang terkait dengan hutan hujan memiliki konektivitas ekologis yang lebih besar daripada yang ada di perkebunan, sehingga berkontribusi pada ekosistem yang lebih terstruktur.
2 BAHAN DAN METODE
2.1 Lokasi penelitian dan pengambilan sampel
Studi kami dilakukan di dataran rendah Provinsi Jambi, Sumatra, Indonesia, dalam rangka proyek EFForTS (Fungsi Ekologi dan Sosial Ekonomi Sistem Transformasi Hutan Hujan Dataran Rendah Tropis). Dua lanskap representatif yang mengalami perubahan ekstensif dalam penggunaan lahan dan berisi area hutan sisa dipilih: Taman Nasional Bukit Duabelas dan Hutan Hujan Harapan (Gambar 1 ). Wilayah ini memiliki iklim tropis lembab dengan dua musim yang berbeda: periode hujan yang berlangsung dari Desember hingga Maret dan periode kemarau yang biasanya terjadi sekitar Juli dan Agustus. Suhu tahunan rata-rata adalah 26,7 °C, dan wilayah ini menerima curah hujan tahunan rata-rata 2235 mm. Wilayah ini telah mengalami konversi hutan hujan yang signifikan menjadi sistem pertanian, terutama untuk membudidayakan tanaman komersial seperti kelapa sawit ( Elaeis guineensis ) dan karet ( Hevea brasiliensis ; Rembold et al., 2017 ). Wilayah studi sebagian besar dicirikan oleh vegetasi alami, khususnya didominasi oleh pohon-pohon yang termasuk dalam famili Dipterocarpaceae (Carneiro de Melo Moura et al., 2022 ).

Plot-plot tersebut terletak di dua lanskap, Bukit Duabelas dan Harapan (yang meliputi THR Sultan Thaha Syalfuddin), di kabupaten Jambi, Sumatra, Indonesia. Plot-plot tersebut mewakili empat jenis penggunaan lahan di wilayah tersebut, yaitu hutan hujan, karet hutan, karet, dan perkebunan kelapa sawit.
Dalam proyek EFForTS, 32 plot berukuran 50 × 50 m di empat tipe penggunaan lahan ditetapkan, dengan delapan plot per tipe penggunaan lahan, yaitu hutan hujan tua bekas tebangan, wanatani karet hutan, dan monokultur karet dan kelapa sawit. Pada setiap subplot berukuran 5 × 5 m yang dipilih secara acak yang dipasang di dalam setiap plot, lima inti tanah dengan diameter 0,04 dan kedalaman 0,20 m diperoleh dari keempat sudut subplot, serta satu di bagian tengah pada tahun 2016. Sampel ditempatkan dalam kotak pendingin selama pengambilan sampel lapangan dan kemudian disimpan pada suhu 4°C di Universitas Jambi. Lima inti per subplot dikumpulkan, diayak, dan disortir sebagai akar halus atau kasar menggunakan dua saringan dengan mata jaring 5 dan 10 mm dan dikeringkan beku sebelum isolasi DNA sampel akar, seperti yang dijelaskan dalam Ballauff et al. ( 2021 ).
2.2 Kerja lapangan dan izin ekspor
Izin Penelitian (Surat Izin Penelitian, nomor referensi: 328/SIP/FRP/E5/Dit.KI/IX/2016) diberikan oleh Kementerian Riset, Teknologi, dan Pendidikan Tinggi (Kementrian Riset, Teknologi dan Pendidikan Tinggi, Jakarta, Indonesia). Persetujuan otorisasi untuk kerja lapangan, pengumpulan sampel, pengangkutan sampel dalam negeri, ekspor sampel, dan akses terhadap materi genetik diberikan oleh Pusat Penelitian Biologi Lembaga Ilmu Pengetahuan Indonesia (Lembaga Ilmu Pengetahuan Indonesia, Jakarta, Indonesia), dengan nomor 2781/IPH.1/KS.02.04/IX/2016 dan B-1345/IPH.1/KS.02.04/III/2019.
Selain itu, izin pengambilan sampel, pengangkutan sampel dalam negeri, ekspor dan akses terhadap materi genetik dikeluarkan oleh Kementerian Lingkungan Hidup dan Kehutanan, Direktorat Jenderal Konservasi Sumber Daya Alam dan Ekosistem Republik Indonesia, nomor S.1068/KKH/SDG/KSA.2/11/2016 dan SK.335/KSDAE/SET/KSA.2/7/2019.
Untuk impor, izin diberikan oleh Kamar Pertanian Niedersachsen (Kantor Perlindungan Tanaman, Hannover, Jerman), sebagaimana didokumentasikan dalam Surat Kuasa (nomor referensi: DE-NI-17-032008-61-EC).
2.3 Prosedur molekuler dan bioinformatika
Dalam penelitian ini, kami menggunakan pendekatan DNA metabarcoding pada sampel akar terpilih untuk mengidentifikasi spesies tanaman dan jamur. Namun, dua sampel—satu dari kelapa sawit dan satu dari petak karet—dikeluarkan dari analisis karena jumlahnya tidak mencukupi (mewakili n = 30 petak). Akar halus yang kering dicacah dalam RETSCH Mixer Mill MM 400 pada 30 Hz selama 20–30 detik hingga menjadi bubuk, disimpan dalam tabung 2 mL, dan disimpan dalam nitrogen cair selama pemrosesan. Isolasi DNA dari akar halus dilakukan menggunakan innuPREP Plant DNA Extraction Kit (Innuscreen GmbH) dengan cara mensuspensikan kembali sampel dalam 100 μL air dan menggunakan 400 μL Lysis Solution SLS mengikuti rekomendasi produsen (Innuscreen GmbH) dan mengganti buffer elusi dengan air bebas nuklease untuk langkah terakhir.
Templat DNA akar diperkuat menggunakan daerah rantai besar ribulosa-1,5-bisfosfat karboksilase ( rbcL ) dengan primer rbcL2 5′-TGGCAGCATTYCGAGTAACTC-3′ (Palmieri et al., 2009 ) dan rbcLa-R 5′-GTAAAATCAAGTCCACCRCG-3′ (Kress & Erickson, 2007 ), yang menghasilkan produk PCR dengan panjang sekitar 450 bp. PCR rangkap tiga dilakukan untuk memungkinkan deteksi spesies tanaman dengan konsentrasi DNA rendah. Setiap reaksi mengandung volume akhir 14 μL menggunakan 6,8 μL H 2 O, 1 μL DNA (10–20 ng/μL), 1,5 μL Buffer PCR 10X (dengan 0,8 M Tris–HCl, 0,2 M (NH 4 ) 2 SO 4 ), 1,5 μL MgCl 2 (25 mM), 1 μL dNTP (2,5 mM setiap dNTP), 1 μL setiap primer maju dan mundur (5 pmol/μL) dan 0,2 μL Taq Hot FirePol (5 U/μL) dari Solis BioDyne (Estonia). Kontrol positif dan negatif dijalankan bersamaan dengan setiap proses PCR, dan tidak ada produk yang diamati dalam sampel kontrol negatif. Pengaturan siklus termal diprogram ke langkah denaturasi awal pada suhu 95°C selama 15 menit, diikuti oleh 35 siklus pada suhu 94°C selama 1 menit, 50°C selama 1 menit, 72°C selama 1 menit, dan langkah ekstensi akhir pada suhu 72°C selama 20 menit. Produk PCR dimurnikan menggunakan GENECLEAN Kit (MP Biomedicals) dan digabungkan sebelum persiapan pustaka. Konsentrasi diukur menggunakan spektrofotometer fluoresensi Qubit (Life Technologies) dan distandarisasi hingga 200 ng per sampel. Illumina TruSeq Nano DNA High Throughput Library Prep Kit (96 sampel) dan Illumina TruSeq DNA CD Indexes digunakan untuk menyiapkan pustaka sekuensing. Pengurutan ujung berpasangan dilakukan pada Illumina MiSeq menggunakan siklus kit MiSeq Reagent v2-300 dengan pustaka pengurutan DNA pada 10 pM dengan kontrol PhiX 10% di NIG (Unit Inti Genomik Integratif NGS di Universitas Göttingen).
Data komunitas jamur yang diidentifikasi menggunakan wilayah ITS diperoleh dari Ballauff dkk. ( 2021 ), yang merupakan bagian dari kumpulan data dari lokasi studi yang sama. Wilayah primer ITS1-F_KYO2 5′-TTYRCTRCGTTCTTCATC-3′ (Toju dkk., 2012 ) dan ITS2 5′-GCTGCGTTCTTCATCGATGC-3′ (White dkk., 1990 ) termasuk adaptor Illumina yang menjorok, digunakan untuk amplifikasi wilayah ITS 1 dan 2 yang menargetkan komunitas jamur. Untuk setiap ekstraksi DNA yang diperoleh dari sampel akar halus, PCR dilakukan menggunakan 0,25 μL Phusion High-Fidelity DNA Polymerase (2 U μL −1 ), 5 μL buffer GC Phusion 5x (Thermo Fisher Scientific, Waltham, AS), 0,075 μL MgCl 2 (50 mM), 1,25 μL DMSO (5%), 1,25 μL bovine serum albumin (8 mg mL −1 ), 0,5 μL campuran dNTP (masing-masing 10 mM, Thermo Fisher Scientific, Osterode am Harz, Jerman), 0,5 μL setiap primer (10 mmol/L, Microsynth, Wolfurt, Austria) dan 2 μL DNA cetakan (5 ng μL −1 ). Campuran PCR disesuaikan dengan volume total 25 μL dengan air bebas dNTP. Parameter siklusnya adalah 1 siklus 98°C selama 30 detik, 30 siklus 98°C selama 10 detik, 47°C selama 20 detik, dan 72°C selama 20 detik, dan ekstensi akhir pada 72°C selama 5 menit. Tiga reaksi replikasi teknis dilakukan dan digabungkan sebelum pengurutan.
Kontrol kualitas pembacaan mentah Illumina yang diperoleh dalam studi ini dilakukan menggunakan FastQC (Andrew, 2010 ). Urutan dipangkas untuk menghilangkan primer dan urutan adaptor menggunakan Cutadapt (Martin, 2011 ). Kami menggunakan Usearch 11.0.667 (Edgar, 2010 ) untuk menggabungkan pembacaan maju dan mundur menggunakan perintah -fastq_mergepairs. Urutan berkualitas rendah (< skor Q 20, <100 bp, pasangan basa ambigu) dan singleton dihilangkan menggunakan perintah -fastq_filter dan -minsize. Pembacaan urutan dide-replikasi (-fastx_uniques), diurutkan berdasarkan ukuran (-sortbysize), dan dikelompokkan (-cluster_otus) menggunakan algoritma UPARSE-OTU (Edgar, 2013 ).
Kami memilih untuk menggunakan Operational Taxonomic Units (OTUs) untuk mengurangi gangguan dan meminimalkan dampak kesalahan sekuensing (Edgar, 2013 ). Semua sekuens OTU kemudian dibandingkan dengan basis data NCBI Genbank untuk memverifikasi keakuratan penugasan OTU menggunakan pencarian Blastn, dengan mengasumsikan leluhur umum terendah yang memiliki nilai kesamaan sekuens tertinggi sebagai penugasan yang benar (Carneiro de Melo Moura et al., 2022 ) dan menggunakan 97% sebagai tingkat kesamaan, yang memungkinkan penugasan setidaknya ke tingkat genus. Berdasarkan literatur yang ada, OTU akar tanaman dikategorikan sebagai asli atau non-asli. Selain itu, mereka diklasifikasikan berdasarkan bentuk kehidupan mereka, seperti pohon, semak atau herba (Kleunen et al., 2019 ; Rembold et al., 2017 ; http://www.plantsoftheworldonline.org/ ).
OTU jamur dikategorikan berdasarkan mode trofik sebagai jamur saprofitik, patotropik, dan simbiotropik dengan menggunakan alat anotasi FUNGuild ( https://github.com/UMNFuN/FUNGuild ; Nguyen et al., 2016 ), sedangkan urutan yang tidak ditetapkan diberi label sebagai tidak teridentifikasi.
Sebanyak 1.916.098 pembacaan ujung berpasangan diperoleh menggunakan uji rbcL untuk akar, berkisar antara 35.948 hingga 91.265 sekuens per sampel (Carneiro de Melo Moura, 2025a , 2025b ). Wilayah ITS yang menargetkan jamur memperoleh total 1.013.513 pembacaan ujung berpasangan, berkisar antara 5762 hingga 76.331 pembacaan per sampel (Carneiro de Melo Moura, 2025c ).
2.4 Analisis hilir
Semua visualisasi data dan analisis data dilakukan menggunakan hitungan sekuens OTU yang dijernihkan untuk akar dan jamur secara terpisah berdasarkan kedalaman penghalusan menggunakan kedalaman sampel minimum dalam paket Phyloseq (McMurdie & Holmes, 2013 ) di RStudio, R versi 4.2.2 (Tim Inti R, 2022 ). Perbedaan antar sampel dihitung menggunakan pohon UPGMA tertimbang berdasarkan jarak UniFrac antara sampel dengan fungsi Unifrac dalam paket R phyloseq (McMurdie & Holmes, 2013 ). Kami menguji perbedaan dalam komunitas akar dan jamur yang terdeteksi dalam empat jenis penggunaan lahan menggunakan Analisis Varians Multivariat Permutasi (PERMANOVA) yang dihitung dengan perbandingan berpasangan jarak Unifrac dalam paket R vegan (Oksanen, 2013 ). Untuk menilai keberadaan autokorelasi spasial dalam keanekaragaman jamur dan akar dalam setiap jenis penggunaan lahan, kami menggunakan statistik Moran’s I. Secara khusus, statistik Moran’s I mengevaluasi apakah distribusi spasial keanekaragaman lebih terkelompok daripada yang diharapkan secara kebetulan. Analisis ini dilakukan menggunakan paket R spdep (Bivand, 2022 ). Moran’s I dihitung untuk berbagai jenis penggunaan lahan dengan membuat matriks bobot spasial berdasarkan tetangga terdekat, lalu menguji pengelompokan spasial yang signifikan dalam data keanekaragaman.
Untuk menentukan hubungan antara penggunaan lahan dan komposisi spesies, kami mengidentifikasi taksa akar dan jamur yang terkait dengan satu atau beberapa jenis penggunaan lahan. Ini dicapai dengan mengimplementasikan fungsi IndVal.g dari paket R indicspecies (De Caceres & Jansen, 2010 ) menggunakan 10 4 permutasi. OTU ditetapkan sebagai indikator untuk jenis penggunaan lahan tertentu berdasarkan koefisien korelasi titik-biserial ( r ), dengan mempertimbangkan hasil yang signifikan pada p < 0,05. Pendekatan ini memperkirakan koefisien korelasi ( r ) dari asosiasi positif OTU dengan satu atau beberapa jenis penggunaan lahan. Indeks IndVal menghitung nilai asosiasi tertinggi antara spesies dan setiap kelompok situs atau kombinasi kelompok situs (De Cáceres et al., 2010 ). Untuk memvisualisasikan komposisi asosiasi OTU yang signifikan ( p < 0,05) (taksa indikator) untuk setiap jenis penggunaan lahan, jaringan bipartit untuk setiap kelompok taksonomi (tumbuhan dan jamur) digunakan menggunakan tata letak Fruchterman–Reingold dengan 10 4 permutasi dalam paket R igraph (Csardi & Nepusz, 2006 ). Lebih jauh , kami menguji kelimpahan OTU diferensial di seluruh penggunaan lahan menggunakan uji rasio kemungkinan (likelihood ratio tests/LRT) dalam paket edgeR (Robinson et al., 2010 ), mengikuti Hartman et al .
Kami membangun dan memvisualisasikan asosiasi antara OTU akar dan jamur dan jenis penggunaan lahan, dengan fokus pada spesies indikator tanaman dan jamur. Untuk melakukannya, jaringan tanaman-jamur dihitung menggunakan korelasi signifikan, nilai- p <0,001 dan koefisien korelasi Spearman ( ρ ) > 0,7, antara OTU akar dan OTU jamur menggunakan 9999 permutasi dan diplot menggunakan algoritma tata letak Fruchterman-Reingold dalam igraph (Csardi & Nepusz, 2006 ). Metrik jaringan, termasuk jumlah simpul, tepi, dan konektivitas simpul rata-rata, diperoleh menggunakan igraph (Csardi & Nepusz, 2006 ).
Kami mengkarakterisasi distribusi OTU dalam jaringan dan interaksinya dalam dan antar modul menggunakan analisis modularitas dengan simulasi annealing (Guimerà & Amaral, 2005 ) yang digunakan dalam paket R rnetcarto (Doulcier & Stouffer, 2015 ) menggunakan OTU yang menunjukkan korelasi Spearman yang signifikan ( p < 0,001, ρ > 0,7). Peran node dalam jaringan ditentukan melalui analisis komprehensif tentang hubungan antara koefisien partisipasi ( P ), yang mencerminkan keragaman koneksi di seluruh komunitas jaringan, dan derajat dalam modul (z), yang mengukur konektivitas setiap node dalam modulnya. Penilaian ini, yang dilakukan menggunakan rnetcarto, menghasilkan kategorisasi node ke dalam peran yang berbeda berdasarkan tingkat konektivitas dan partisipasinya. Secara khusus, node diklasifikasikan sebagai kinless ketika tepinya terdistribusi secara merata di antara semua modul ( p > 0,80), konektor jika mereka menunjukkan banyak tepi ke modul lain (0,62 < p ≤ 0,80), peripheral jika mereka menampilkan sebagian besar tautan mereka dalam modul mereka (0,05 < p ≤ 0,62), dan ultra-peripherals jika mereka menyajikan semua tepinya dalam modul mereka ( p ≤ 0,05; Guimerà & Amaral, 2005 ). Klasifikasi ini memberikan pemahaman bernuansa tentang peran setiap node dalam jaringan, menjelaskan pola konektivitasnya dan kontribusinya terhadap keseluruhan struktur jaringan (Guimerà & Amaral, 2005 ).
Kami menguji signifikansi statistik modularitas menggunakan kode netcarto-legacy dari pustaka RGraph ( https://github.com/cran/rnetcarto/tree/master/src/rgraph ) yang ditulis dalam C dan dikembangkan oleh Guimerà dan Amaral ( 2005 ). Kami menjalankan 25 pengacakan di fasilitas klaster High Performance Computing di CSIRO. Keluaran uji pengacakan memberikan rata-rata dan simpangan baku modularitas jaringan acak, yang digunakan untuk menghitung z -score dan nilai p masing-masing menggunakan interval kepercayaan 95%.
3 HASIL
3.1 Gambaran umum data sekuens dan struktur komunitas
Di antara OTU akar yang teridentifikasi, 247 diklasifikasikan ke dalam 25 ordo, 47 famili, 111 genera dan 124 spesies tumbuhan. Penugasan ini terdiri dari 13 OTU yang dikategorikan sebagai spesies herba, 56 sebagai semak dan 178 sebagai spesies pohon. Lebih jauh, penugasan sekuens mengungkapkan keberadaan delapan spesies yang bukan asli Indonesia. Selain itu, set data kami mengidentifikasi 40 OTU pada tingkat genus, yang berpotensi menunjukkan ketersediaan sekuens yang tidak lengkap untuk spesies Indonesia dalam basis data sekuens atau kurangnya perbedaan diagnostik pada tingkat spesies dengan kode batang rbcL . Pohon UPGMA tertimbang mengungkapkan efek penggunaan lahan yang terlihat pada komposisi komunitas akar ( p < 0,005 untuk semua, kecuali antara kelapa sawit dan karet; Gambar S1A ).
Untuk urutan jamur, 14.049 OTU dikategorikan ke dalam 177 ordo, 430 famili, dan 1.159 genus. Dalam OTU yang ditetapkan, 2.010 OTU diklasifikasikan sebagai simbiotrof, 2.362 sebagai saprotrof, 540 sebagai patotrof, dan 1.067 sebagai bagian dari beberapa serikat fungsional. Sebanyak 8.070 OTU jamur tidak dapat ditetapkan ke dalam kelompok fungsional mana pun. Pohon UPGMA tertimbang untuk kumpulan data jamur menunjukkan efek minimal dari perubahan penggunaan lahan pada profil komunitas jamur ( p = 0,709; Gambar S1B ).
Uji autokorelasi spasial Moran ‘s I menggunakan keragaman akar Shannon di berbagai jenis penggunaan lahan menunjukkan distribusi non-acak di hutan hujan, dengan statistik Moran’s I sebesar 0,1547 (SD = 2,9804, p = 0,00144), yang menunjukkan pengelompokan keragaman spasial yang signifikan ( p < 0,01). Sebaliknya, pola spasial dalam penggunaan lahan yang dikonversi konsisten dengan distribusi acak (karet hutan: Moran’s I = −0,218, SD = −0,9823, p = 0,837; kelapa sawit: Moran’s I = 0,1941, SD = −0,4624, p = 0,6781; dan karet: −0,1989, SD = −0,5583, p = 0,7117).
Demikian pula, autokorelasi spasial keanekaragaman jamur di berbagai jenis penggunaan lahan menunjukkan kecenderungan yang sedikit signifikan terhadap pengelompokan spasial di hutan hujan, dengan statistik Moran’s I sebesar -0,0075 (SD = 1,3545, p = 0,08778). Sebaliknya, pola spasial di jenis penggunaan lahan lainnya konsisten dengan distribusi acak (karet hutan: Moran’s I = -0,1792, p = 0,6742; kelapa sawit: Moran’s I = -0,1985, p = 0,6837; dan karet: Moran’s I = -0,2239, p = 0,8425).
3.2 Jaringan bipartit spesies indikator yang terkait dengan penggunaan lahan
Keanekaragaman spesies indikator tanaman dan kelompok fungsional menunjukkan variasi di antara tipe penggunaan lahan (Gambar 2 , Lampiran S1 ). Hutan hujan menunjukkan keanekaragaman spesies indikator yang lebih tinggi, meliputi susunan taksa asli dan endemik yang lebih besar, serta bentuk kehidupan tanaman yang beragam (Gambar 2a,b ). Sebaliknya, sistem monokultur yang lebih intensif mengungkapkan keberadaan spesies tanaman non-asli seperti Asystasia gangetica dan Clidemia hirta , bersama dengan peningkatan jumlah spesies herbal lainnya (Gambar 2c ). Perbedaan komposisi terlihat jelas di antara tipe penggunaan lahan untuk komunitas akar dan jamur, dengan kesamaan yang lebih tinggi diamati antara tipe penggunaan lahan yang kurang intensif, seperti hutan hujan dan karet hutan (Gambar 2a ). Sebaliknya, tipe penggunaan lahan yang lebih intensif, seperti monokultur, menampilkan lebih sedikit spesies yang diidentifikasi sebagai indikator. Spesies indikator pada plot perkebunan kelapa sawit meliputi Elaeis guineensis (pohon kelapa sawit), bersama spesies indikator lain seperti spesies herba Centotheca lappacea dan herba merambat non-asli Asystasia gangetica dan Clidemia hirta . Pada monokultur karet, spesies indikator meliputi Hevea brasiliensis (pohon karet), semak merambat asli Ichnocarpus frutescens , Trachelospermum asiaticum , dan herba C. lappacea , dan C. hirta .

Jaringan bipartit spesies indikator yang terkait dengan empat jenis penggunaan lahan: Hutan hujan, karet hutan, karet, dan monokultur kelapa sawit. (a) Jaringan spesies indikator tanaman pada tingkat ordo. (b) Kelimpahan relatif akar tanaman indikator pada empat jenis penggunaan lahan. (c) Kelimpahan relatif bentuk kehidupan taksa tanaman indikator pada empat jenis penggunaan lahan. (d) Jaringan bipartit spesies indikator jamur yang diidentifikasi dalam sampel akar pada tingkat filum. (e) Kelimpahan relatif taksa jamur indikator pada empat jenis penggunaan lahan. (f) Kelimpahan relatif serikat fungsional taksa jamur indikator pada empat jenis penggunaan lahan. Warna yang digunakan dalam gambar sesuai dengan masing-masing penugasan Unit Taksonomi Operasional (OTU).
Perubahan penggunaan lahan memengaruhi kelompok taksonomi dan fungsional yang terkait dengan spesies indikator jamur, karena komposisi dan proporsi spesies indikator bervariasi di sepanjang penggunaan lahan (Gambar 2 ). Jaringan jamur menggambarkan hubungan yang erat antara hutan hujan dan klaster karet hutan, sementara monokultur menunjukkan kedekatan yang lebih besar satu sama lain (Gambar 2d ). Hutan hujan menunjukkan indikator jamur yang berbeda dari filum Calcarisporiellomycota dan Kickxellomycota. Sebaliknya, OTU yang ditugaskan ke Mucoromycota dan Mortierellomycota lazim sebagai kelompok indikator dalam monokultur (Gambar 2e ). Sebagian besar pembacaan yang terkait dengan monokultur di antara taksa indikator jamur termasuk patogen tanaman spesifik spesies potensial, sementara AMF yang diidentifikasi sebagai OTU indikator lebih melimpah di lingkungan yang kurang terganggu (Gambar 2f ).
3.3 Jaringan ko-kemunculan antara akar dan jamur
Analisis jaringan mengungkapkan bahwa modularitas, yaitu partisi atau klaster dengan koneksi yang lebih rapat, dan konektivitas, yaitu tingkat di mana simpul saling terhubung, dipengaruhi oleh jenis penggunaan lahan dan kelompok fungsional tanaman. Jaringan tanaman-jamur terdiri dari 2296 simpul (163 OTU akar dan 2133 OTU jamur), dan 47.922 tepi (42.217 jamur-jamur, 580 tanaman-tanaman, dan 5125 tanaman-jamur) (Gambar 3a , Lampiran S2 ). Modularitas tinggi ( Q = 0,9) menunjukkan struktur komunitas yang kuat dalam jaringan, dengan simulasi annealing mengoptimalkan partisi menjadi 22 modul. Uji pengacakan modularitas menunjukkan bahwa jaringan tersebut secara signifikan modular ( z -score = 37,504, p -value <0,0001). Rata-rata modularitas jaringan acak adalah Mrand = 0,2891 dan simpangan baku 0,0019. Konektivitas tertinggi diamati dalam modul, yang mencakup spesies indikator yang terkait dengan hutan hujan, dengan 16.418 tepi, yaitu asosiasi. Sebaliknya, spesies indikator karet hutan memiliki 2981 tepi, kelapa sawit 2883 tepi dan karet 203 tepi. Ada lebih sedikit asosiasi tumbuhan-jamur yang signifikan ketika mempertimbangkan OTU akar yang ditugaskan untuk spesies tumbuhan herba (194), berbeda dengan jumlah asosiasi tumbuhan-jamur yang lebih besar yang diamati untuk pohon (3617) atau semak (1025) (Data S1 ). Node terputus yang diamati dalam jaringan (yaitu Kunstleria ridleyi, Poikilospermum suaveolens dan Suregada glomerulata ) dapat menunjukkan bahwa spesies ini sangat terspesialisasi, tidak memiliki interaksi ekologi langsung, atau bahwa beberapa interaksi tidak terdeteksi karena data yang tidak lengkap (Gambar 3a ).
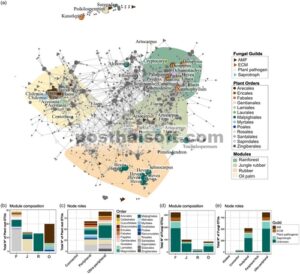
(a) Jaringan berdasarkan korelasi ko-kemunculan yang signifikan antara akar tanaman dan jamur. Modul yang ditampilkan menyoroti klaster yang berisi OTU tanaman indikator yang terkait dengan empat jenis penggunaan lahan (hutan hujan, karet hutan, karet, dan perkebunan kelapa sawit), menggunakan analisis modularitas dengan simulasi anil. Lingkaran menunjukkan penugasan OTU tanaman indikator yang disajikan pada tingkat genus; warna menggambarkan urutan OTU indikator tanaman. Segitiga menggambarkan kelompok indikator jamur yang diwarnai oleh serikat fungsional (AMF, EMF, patogen tanaman, dan saprotrof). (b) Komposisi tanaman untuk modul yang mencakup OTU tanaman indikator, dan (c) profil taksonomi peran simpul yang diidentifikasi melalui penugasan OTU tanaman yang dikelompokkan dalam modul interkingdom. (d) Komposisi jamur untuk modul yang disorot yang berisi OTU jamur indikator. (e) Profil taksonomi peran simpul yang diidentifikasi melalui penugasan serikat jamur yang dikelompokkan dalam modul interkingdom. Warna sesuai dengan penugasan OTU akar tanaman pada tingkat ordo dan jamur pada tingkat serikat. N ° = Nomor.
3.4 Peran topologi jaringan
Jaringan ini terutama terdiri dari simpul yang dikategorikan sebagai perifer, dengan 68 OTU tanaman dan 712 OTU jamur, atau ultra-perifer, dengan 94 OTU tanaman dan 1327 OTU jamur (Gambar 3a ). Selain itu, 73 simpul (satu akar dan 72 OTU jamur) diidentifikasi sebagai konektor (Gambar 3 dan 4 , Lampiran S1 ). Gambar 3b–e mengilustrasikan komposisi taksonomi modul yang terkait dengan spesies indikator yang terkait dengan empat jenis penggunaan lahan. Di antara modul-modul ini, spesies indikator hutan hujan menunjukkan keanekaragaman dan kelimpahan OTU tertinggi (Gambar 3b ). Selain itu, modul-modul yang terkait dengan hutan hujan ini menunjukkan proporsi terbesar AMF, EMF, dan jamur saprofit (Gambar 3d ). Spesies yang menempati peran perifer dan ultra-perifer mencakup berbagai kelompok taksonomi atau peran fungsional, seperti yang diilustrasikan dalam Gambar 3c,e .

Peran simpul topologi berdasarkan distribusi derajat modul (z) dan koefisien partisipasi ( P ) dari setiap OTU. (a) Simbol melingkar mewakili OTU tumbuhan, dan (b) simbol segitiga mewakili OTU jamur. Warna sesuai dengan peran simpul. Ambang batas setiap kategori simpul diwakili oleh garis yang dilacak. Derajat dalam modul z[i] mengukur tingkat konektivitas setiap simpul [i] dalam modul yang sama. Nilai z[i] yang lebih tinggi menunjukkan koneksi dalam modul yang lebih kuat. Koefisien partisipasi mengukur distribusi tautan simpul [i] di berbagai modul; mendekati 1 jika tautan didistribusikan secara merata di semua modul dan 0 jika semua tautan tetap terkurung dalam modulnya sendiri. Simpul dengan z -score ≥2,5 dikategorikan sebagai hub modul, sedangkan simpul dengan z -score <2,5 diklasifikasikan sebagai non-hub.
Jamur Dendrosporium sp. dan Gerhardtia highlandensis diklasifikasikan sebagai penghubung dan indikator sistem hutan hujan. Glomeraceae sp. dan Herpotrichiellaceae sp. merupakan indikator sistem karet hutan dan juga berperan sebagai penghubung. Taksa-taksa ini terkait dengan lingkungan yang menunjukkan tingkat gangguan yang berbeda-beda. Pada kelapa sawit dan karet, penghubung dan indikator meliputi jamur saprotrofik atau taksa yang terkait dengan kondisi oligotrofik, seperti Ascomycota sp., Metschnikowia spp., Pezizales sp., Setophoma terrestris , Herpotrichiellaceae sp., Conlarium sp., Sordariomycetes sp., Ochroconis spp., Cladophialophora spp., Pezizomycotina sp. dan Glomeromycota sp.
Dalam jaringan tersebut, hanya tiga simpul yang diklasifikasikan sebagai kinless karena konektivitasnya yang tinggi. Dua simpul diidentifikasi sebagai EMF: Pachyphlodes melanoxantha dan Elaphomyces favosus . Simpul ketiga termasuk dalam kategori jamur yang tidak diketahui. Perlu dicatat bahwa keragaman jamur indikator yang menempati peran perifer atau ultra-perifer sangat luas dan mencakup berbagai kelompok fungsional (Lampiran S2 dan Data S2 ; Gambar 4 ).
4 DISKUSI
4.1 Pola dalam komunitas tumbuhan-jamur
Temuan kami sejalan dengan gagasan bahwa perakitan komunitas jamur akar dipengaruhi oleh perubahan penggunaan lahan (Ballauff et al., 2020 , 2021 ). Misalnya, penelitian sebelumnya telah menunjukkan bahwa alih fungsi hutan hujan mengakibatkan perubahan substansial dalam komposisi komunitas jamur tanah, meskipun tidak diamati adanya penurunan keanekaragaman yang signifikan (Ballauff et al., 2021 ; Brinkmann et al., 2019 ). Dampak praktik penggunaan lahan intensif dan modifikasi filter abiotik dan biotik yang diakibatkannya di mikrohabitat merupakan efek warisan penggunaan lahan sebelumnya terhadap komposisi komunitas mikrobioma (Ballauff et al., 2020 ; Fichtner et al., 2014 ; Romdhane et al., 2022 ). Pengurangan dalam keanekaragaman akar dapat menyebabkan pengurangan dalam ketersediaan sumber daya dan fungsi ekosistem, karena banyak spesies mungkin bergantung pada jenis akar tertentu untuk pertumbuhan dan kelangsungan hidup mereka (Montesinos-Navarro et al., 2019 ; Põlme et al., 2018 ). Lebih jauh, gangguan interaksi ekologis seperti asosiasi tumbuhan-jamur memengaruhi penyerapan nutrisi dan pertumbuhan tanaman, yang pada gilirannya dapat memengaruhi herbivora, penyerbuk, dan spesies lainnya (Hazard et al., 2013 ; Mommer et al., 2018 ; Scherber et al., 2010 ).
Perbedaan dalam distribusi spasial keanekaragaman tumbuhan dan jamur antara hutan hujan dan penggunaan lahan yang dikonversi mencerminkan bahwa di hutan hujan, spesies langka dan endemik lebih berkelompok secara spasial daripada terdistribusi secara acak. Distribusi keanekaragaman yang tidak acak menunjukkan ekosistem yang sehat, kaya akan heterogenitas habitat dan interaksi spesies (Sritharan et al., 2021 ). Sebaliknya, habitat yang homogen, seperti yang diamati pada penggunaan lahan yang dikonversi, menunjukkan penyederhanaan habitat dan hilangnya keanekaragaman relung dan proses ekologi. Mempertimbangkan distribusi yang tidak acak di hutan hujan sangat penting untuk memahami tidak hanya dampak konversi hutan tetapi juga konservasi dan restorasi (Montoya-Sánchez et al., 2023 ).
111 genus tanaman yang dideteksi dalam studi ini menggunakan DNA metabarcoding mencakup sekitar 20% dari genus yang sebelumnya didokumentasikan di area yang sama menggunakan inventaris tanaman berbasis morfologi yang dilakukan oleh konsorsium penelitian kami selama periode pengambilan sampel 18 bulan (Rembold et al., 2017 ). Perlu dicatat bahwa penelitian yang hanya berfokus pada spesimen individu akar halus yang dilakukan di hutan tropis atau eksperimen keanekaragaman hayati menghadapi tantangan dalam menangkap jumlah kekayaan OTU tanaman yang sama seperti yang diamati dalam penelitian kami (Edy et al., 2022 ; Francioli et al., 2021 ; Gui et al., 2017 ; Koorem et al., 2017 ). Oleh karena itu, cakupan yang lebih lengkap dari keragaman akar dan interaksi menggunakan DNA metabarcoding di wilayah yang sangat beragam di bawah tekanan perubahan global saat ini mungkin mendapat manfaat dari peningkatan jumlah sampel. Meskipun 20% mungkin tampak rendah dalam hal menangkap keragaman genera secara menyeluruh, penting untuk menyadari bahwa metode yang digunakan dalam studi kami memberikan gambaran singkat tentang profil lanskap. Ini dapat berharga untuk penilaian cepat tren umum dalam keragaman tanaman. Namun, kami mengakui bahwa meningkatkan jumlah inti tanah atau pengambilan sampel kemungkinan akan menghasilkan representasi yang lebih komprehensif tentang komposisi spesies di area tersebut. Pendekatan ini berfungsi sebagai tinjauan awal, dengan potensi untuk studi yang lebih rinci menyusul.
4.2 Struktur jaringan akar jamur
Tingkat kompartementalisasi jaringan yang tinggi yang diamati dalam analisis jaringan kami menunjukkan hubungan spesifik antara perakitan komunitas akar dan jamur (Grilli et al., 2016 ; Põlme et al., 2018 ). Penurunan spesies inang dalam monokultur sering dikaitkan dengan kemunculan spesies yang toleran terhadap stres dan oportunistik (Gioria et al., 2023 ; Nayanakantha, 2007 ; Rembold et al., 2017 ), yang dalam penelitian kami diwakili oleh spesies oportunistik dari kedua tanaman (misalnya C. hirta dan A. gangetica ) dan jamur ( Cladophialophora sp.). Spesies generalis ini tumbuh subur dalam kondisi sumber daya rendah (Coleine et al., 2022 ; Robinson & Strauss, 2020 ), yang mendukung bahwa spesies generalis atau toleran stres juga dapat bertindak sebagai penghubung, memfasilitasi interaksi dalam lingkungan yang menantang (Grilli et al., 2016 ; Robinson & Strauss, 2020 ). Lebih jauh lagi, AMF menunjukkan koneksi yang relatif sedikit ke simpul kelapa sawit, yang menunjukkan bahwa AMF lebih terspesialisasi dalam jaringan tersebut. Hal ini mendukung bukti yang berkembang bahwa praktik penggunaan lahan intensif dalam monokultur kelapa sawit dapat berdampak negatif pada keanekaragaman AMF (Edy et al., 2022 ; Phosri et al., 2010 ).
Perubahan lingkungan global pasti mengganggu interaksi biotik, mendorong keberhasilan pembentukan dan penyebaran kelompok taksonomi non-asli (DeWalt et al., 2004 ; Gioria et al., 2023 ; Hulme, 2017 ; Richardson & Rejmánek, 2011 ; Walther et al., 2009 ). Proses atau pendorong utama invasi yang berhasil mencakup gangguan karena fluks sumber daya fisik atau kimia, seperti iklim dan pemupukan (Gioria et al., 2023 ; Walther et al., 2009 ). Penggerak eksternal ini bertindak sebagai filter lingkungan, yang mendukung spesies yang mudah beradaptasi dan oportunistik yang menempati rentang relung ekologi yang lebih luas, yang pada akhirnya mengalahkan spesies penghuni (Gioria et al., 2023 ). Spesies non-asli memiliki karakteristik khusus yang meningkatkan ketahanan dan dapat mendukung perekrutan bibit yang berhasil (DeWalt et al., 2004 ; Richardson & Rejmánek, 2011 ). Sifat-sifat seperti penyebaran jarak jauh, plastisitas reproduksi, kemampuan fiksasi nitrogen, dan alelopati merupakan keunggulan kompetitif dibandingkan spesies asli yang meningkatkan persistensi benih bahkan dalam kondisi yang tidak menguntungkan (DeWalt et al., 2004 ; Gioria et al., 2023 ; Hulme, 2017 ; Richardson & Rejmánek, 2011 ; Walther et al., 2009 ). Akibatnya, interferensi ini memberi mereka keunggulan kompetitif dibandingkan spesies asli yang bergantung pada hubungan mutualistik. Misalnya, semak menahun invasif C. hirta , yang diidentifikasi dalam studi ini sebagai spesies indikator monokultur kelapa sawit, memiliki laju pertumbuhan yang cepat, produksi biji yang produktif, toleransi terhadap naungan, dan melepaskan alelokimia ke dalam tanah yang memberinya keunggulan kompetitif atas flora asli (DeWalt et al., 2004 ). Selain itu, spesies ini toleran terhadap kondisi lingkungan yang ekstrem, menghuni daerah yang terganggu, dan tumbuh subur di habitat terbuka yang dimodifikasi seperti pinggir jalan dan perkebunan pohon (DeWalt et al., 2004 ; Hartman et al., 2018 ; Rembold et al., 2017 ; Richardson & Rejmánek, 2011 ). Lebih jauh lagi, herba merambat non-asli A. gangetica telah dilaporkan memberikan dampak yang merugikan pada hasil perkebunan kelapa sawit. Ia menunjukkan penyebaran yang cepat dan ekspansif dan memiliki sifat generalis yang memungkinkannya beradaptasi dengan berbagai jenis tanah. Kedua tanaman herbal tersebut bersaing dengan tanaman pangan dan spesies asli untuk mendapatkan sumber daya (Kustyanti & Horne, 1991 ; Wibawa et al., 2009).). Lebih jauh lagi, keberadaan spesies tanaman non-asli yang oportunistik dapat memengaruhi keanekaragaman mikrobiota tanah dengan mendorong perubahan tanah melalui proses enzimatik dan efek langsung atau tidak langsung pada siklus karbon dan nitrogen (Gioria et al., 2023 ).
4.3 Dampak penggunaan lahan terhadap konektivitas jaringan
Spesies indikator dari sistem yang sehat atau kurang terkonversi telah menunjukkan konektivitas yang lebih tinggi dalam jaringan (Brinkmann et al., 2019 ), yang menggarisbawahi pentingnya menilai mikrobiota yang terkait dengan akar. Selain itu, taksa indikator yang terkait dengan agroekosistem karet hutan, yang berperan sebagai zona transisi antara habitat yang terganggu ringan dan terganggu, mencakup taksa indikator yang terkait dengan kedua jenis habitat. Misalnya, Glomeraceae spp. dan Herpotrichiellaceae spp. diidentifikasi sebagai indikator karet hutan, dengan AMF dan kelompok tahan stres memainkan peran penting dalam membentuk jaringan (Francioli et al., 2021 ; Hofstetter et al., 2014 ; van der Heijden & Hartmann, 2016 ). Spesies tumbuhan dan jamur yang diidentifikasi sebagai spesies indikator hutan hujan kemungkinan memberikan pengaruh besar pada aliran sumber daya. Akibatnya, penghilangan spesies ini dapat mengakibatkan perubahan signifikan pada komposisi komunitas dan fungsi ekosistem (Guimerà & Amaral, 2005 ). Dalam studi ini, spesies indikator pohon yang diidentifikasi dari hutan hujan termasuk dalam ordo Ericales, Gentianales, Laurales, Magnoliales, Malpighiales, Myrtales, Santalales, dan Sapindales. Mengidentifikasi berbagai spesies indikator yang terkait dengan jenis penggunaan lahan tertentu memberikan wawasan berharga untuk upaya restorasi dan dapat dieksplorasi lebih lanjut melalui eksperimen pengayaan keanekaragaman hayati (Paterno et al., 2024 ).
Pengelolaan konservasi dan pemulihan fungsi dan jasa ekosistem dapat dicapai dengan berfokus pada jaringan interaksi dan menargetkan spesies indikator guna meningkatkan stabilitas ekosistem (Harvey et al., 2017 ).
5 KESIMPULAN
Studi kami merupakan upaya perintis untuk mengeksplorasi akar tanaman dan hubungannya dengan komunitas jamur dalam kerangka sistem konversi hutan menggunakan kode batang tanaman dan jamur dari sampel DNA yang identik. Berdasarkan analisis kami, kami telah mengamati koeksistensi antara tanaman pangan yang dibudidayakan dan organisme yang tahan terhadap stres atau generalis. Menariknya, sistem agroforestri dapat berfungsi sebagai zona perantara antara habitat yang terganggu ringan dan sangat terganggu, yang menampung spesies jamur indikator yang merupakan karakteristik dari kedua tipe habitat tersebut.
Kami mengidentifikasi perbedaan topologi dan struktural di antara komunitas jamur dan tumbuhan di berbagai jenis penggunaan lahan. Modul yang terhubung dengan jenis penggunaan lahan yang kurang terkelola menunjukkan konektivitas yang lebih baik dan menampung spesies indikator yang terkait dengan sistem hutan hujan atau karet hutan. Hal ini berbeda dengan modul yang berisi spesies indikator yang terkait dengan plot perkebunan. Selain itu, taksa langka atau endemik sering kali memainkan peran penting dalam struktur jaringan dengan membentuk beberapa koneksi antara modul yang berbagi spesies indikator yang terkait dengan habitat hutan hujan. Hal ini menyoroti dampak monokultur pada komposisi dan struktur jaringan tumbuhan-jamur dan menunjukkan peran potensial sistem agroforestri dalam menegakkan keanekaragaman, koneksi, dan selanjutnya, fungsi ekosistem.
KONTRIBUSI PENULIS
Carina Carneiro de Melo Moura, Valentyna Krashevska, Nathaly Guerrero-Ramírez, Johannes Ballauff, Andrea Polle, dan Oliver Gailing merancang penelitian ini. Carina Carneiro de Melo Moura melakukan analisis bioinformatika, analisis hilir, visualisasi data, analisis statistik, dan menulis draf naskah. Valentyna Krashevska, Nathaly Guerrero-Ramírez, Oliver Gailing, Francisco Encinas-Viso, dan Ulfah J. Siregar berkontribusi dalam penulisan. Johannes Ballauff dan Andrea Polle menyediakan kumpulan data jamur. Semua penulis meninjau draf dan berkontribusi dalam perbaikan artikel ini.
UCAPAN TERIMA KASIH
Kami berterima kasih kepada Gudrun Diederich dan Larissa Kunz atas dukungan teknis untuk eksperimen laboratorium, Oleksandra Dolynska, Thomas Klein, dan Fabian Ludewig atas saran teknis. Kami juga menghargai upaya asisten lapangan yang terlibat dalam pengambilan sampel. Kami juga berterima kasih kepada Nunzio Knerr atas bantuannya dalam menyiapkan simulasi jaringan. Studi ini merupakan bagian dari proyek Collaborative Research Centre 990—EFForTS (Fungsi Ekologis dan Sosial Ekonomi Sistem Transformasi Hutan Hujan Dataran Rendah Tropis, https://www.uni-goettingen.de/efforts ) ID 192626868—SFB 990 (B14 dan B07) dari Yayasan Penelitian Jerman (DFG). Kami berterima kasih kepada Open Access Publication Funds dari University of Göttingen. Kami menghargai kesempatan untuk melakukan analisis pada High-Performance Clusters dari GWDG dan CSIRO. mengakui pendanaan DFG untuk Biodiversa+ BioMonI (533271599).
PERNYATAAN KONFLIK KEPENTINGAN
Penulis menyatakan tidak ada benturan kepentingan.