
ABSTRAK
Toleransi kekeringan spesies pohon menjadi perhatian dalam konteks perubahan iklim, dan analisis lingkaran pohon dapat digunakan untuk menilai respons pertumbuhan masa lalu, terhadap peristiwa kekeringan. Dalam studi terkini, kami menerapkan pendekatan ini pada 1281 individu dengan silsilah yang diketahui di perkebunan uji genetik jangka panjang pinus lodgepole di Kanada bagian barat. Kami menilai metrik resistensi, ketahanan, dan pemulihan, dan menganalisis hubungan kausalnya dengan pertumbuhan jangka panjang dan kerentanan terhadap penyakit melalui pemodelan persamaan struktural. Kami menemukan bahwa pohon dengan ketahanan jangka pendek yang rendah terhadap peristiwa kekeringan juga mengalami pengurangan yang parah dalam pertumbuhan jangka panjang. Heritabilitas dalam arti sempit dari metrik toleransi kekeringan rendah untuk respons jangka pendek di lokasi tertentu, sementara indeks penurunan jangka panjang baru untuk famili menunjukkan heritabilitas sedang (mathematical equationdari 0,15 hingga 0,30 ± 0,03). Kami juga mendeteksi bukti adaptasi lokal, dengan pohon-pohon dari dataran rendah menunjukkan adaptasi kekeringan yang lebih baik. Kami menyimpulkan bahwa pemilihan genotipe untuk toleransi kekeringan adalah mungkin, dan bahwa spesies atau populasi lain dapat disaring menggunakan metode ini. Kami juga mencatat bahwa indeks penurunan jangka panjang baru yang dikembangkan dalam penelitian ini menunjukkan tingkat kontrol genetik yang lebih tinggi daripada metrik lainnya, dan karena itu mungkin lebih menarik dalam penelitian dendrokronologi.
1 Pendahuluan
Perubahan iklim diperkirakan akan menyebabkan peningkatan suhu rata-rata yang stabil di wilayah boreal dan sedang, dikombinasikan dengan peningkatan frekuensi dan tingkat keparahan peristiwa kekeringan ekstrem (IPCC 2014 ). Perubahan kondisi iklim ini dapat melampaui kapasitas adaptif spesies pohon, mengurangi kekuatan pohon, meningkatkan kerentanan terhadap stresor biotik dan abiotik, dan menyebabkan penurunan populasi karena mortalitas yang meningkat secara signifikan (Martínez-Vilalta et al. 2012 ; McDowell et al. 2008 ). Selama beberapa dekade terakhir, stres kekeringan telah dikaitkan dengan kematian pohon yang meluas, pertumbuhan yang berkurang dan penyerapan karbon, baik di hutan boreal maupun sedang (Allen et al. 2010 ; Hogg et al. 2017 ; Wong dan Daniels 2017 ).
Pohon telah mengembangkan mekanisme adaptasi kekeringan yang berbeda, yang sering diklasifikasikan sebagai strategi toleransi kekeringan dan penghindaran kekeringan (Oliveira et al. 2021 ). Mekanisme toleransi kekeringan melibatkan pengembangan jaringan xilem yang tahan kekeringan, yang mampu menahan potensial air rendah dan menghindari kavitasi (Feng et al. 2021 ). Secara anatomi, hal ini sebagian besar disebabkan oleh produksi kayu yang lebih padat, dan trakeid dengan lumen yang lebih kecil dan dinding sel yang lebih tebal, yang dapat mengurangi potensial air minimum di mana xilem mulai mengalami kegagalan hidrolik (Chave et al. 2009 ; Hacke et al. 2001 ). Mekanisme penghindaran kekeringan, di sisi lain, bergantung pada pemeliharaan potensial air yang lebih tinggi di xilem melalui penutupan stomata awal dan sintesis senyawa osmoprotektif, atau menginvestasikan sumber daya karbon ke dalam produksi akar untuk penyerapan air yang lebih efisien (McDowell et al. 2008 ). Respons plastik dapat meningkatkan ketahanan pohon terhadap kekeringan dengan mengandalkan mekanisme ini, sementara bukti lain menunjukkan bahwa plastisitas fenotipik individu tidak cukup untuk mengatasi peningkatan stres kekeringan yang terkait dengan perubahan iklim yang cepat di ekosistem hutan dengan keterbatasan kelembaban (Anderegg et al. 2019 ). Studi adaptasi kekeringan pada konifer telah menunjukkan perbedaan genetik yang kuat antara populasi, yang telah dikaitkan dengan adaptasi lokal (George et al. 2017 ; Housset et al. 2018 ; Isaac-Renton et al. 2018 ) dan divergensi genetik (George et al. 2017 ). Kontrol genetik sifat-sifat yang beradaptasi dengan kekeringan menawarkan kemungkinan untuk memilih genotipe spesifik yang menghasilkan hutan yang lebih produktif dan tangguh.
Bahasa Indonesia: Untuk memilih genotipe yang beradaptasi terhadap kekeringan, metrik yang kuat dari adaptasi kekeringan perlu ditetapkan, dan kontrol genetik dari sifat-sifat ini (yaitu, heritabilitasnya) perlu ditentukan dan diverifikasi. Lingkaran pohon menawarkan catatan terperinci dari respons pertumbuhan genotipe individu terhadap kekeringan dan peristiwa iklim dan pengelolaan lainnya (Fritts 1976 ). Analisis lingkaran pohon, atau dendrokronologi, telah menjadi terkenal dalam beberapa tahun terakhir sebagai alat untuk mempelajari respons kekeringan pada banyak spesies pohon (Camarero et al. 2015 ; Gazol et al. 2018 ; Serra-Maluquer et al. 2018 ; Zang et al. 2014 ). Dua atribut menarik yang dapat diekstraksi dari analisis lingkaran pohon adalah, kemampuan untuk mempertahankan pertumbuhan selama peristiwa kekeringan (resistensi), dan kemampuan untuk pulih ke tingkat pertumbuhan sebelum kekeringan setelah peristiwa kekeringan (ketahanan, Lloret et al. 2011 ). Ciri-ciri ini telah digunakan untuk mempelajari adaptasi lokal terhadap kekeringan pada beberapa konifer (Depardieu et al. 2020 ; Isaac-Renton et al. 2018 ; Montwé et al. 2016 ), tetapi pertanyaan tentang bagaimana ketahanan dan resistensi memengaruhi pertumbuhan pohon, setelah kejadian kekeringan, masih belum terjawab. Sementara hubungan antara ketahanan yang rendah dan peningkatan kematian konifer baru-baru ini diamati dalam skala global (DeSoto et al. 2020 ), ada juga hipotesis bahwa pemulihan yang lebih lambat dari stres kekeringan dapat menunjukkan investasi cadangan karbon ke kayu yang lebih tahan, mengorbankan pertumbuhan yang lebih cepat (Gessler et al. 2020 ). Menetapkan hubungan genetik antara respons jangka pendek terhadap kekeringan dan pertumbuhan jangka panjang dapat membantu mengidentifikasi sifat-sifat spesifik yang menyusun pohon yang beradaptasi dengan kekeringan.
Kontrol genetik dari sifat fisiologis pohon dapat dipelajari dalam percobaan kebun umum, seperti uji keturunan. Percobaan ini ditanam dengan bibit yang berasal dari benih yang dikumpulkan dari lokasi yang berbeda dan ditanam di beberapa lokasi pengujian, di mana semuanya tumbuh dalam kondisi lingkungan yang sama. Pengaturan eksperimental ini terutama dimaksudkan untuk mengendalikan efek lingkungan untuk mengungkap perbedaan genetik, mengukur plastisitas fenotipik, dan mengukur interaksi genotipe-oleh-lingkungan (de Villemereuil et al. 2016 ). Sebagian besar studi adaptasi kekeringan, menggunakan dendrokronologi, berfokus pada satu kejadian kekeringan atau satu lokasi (Depardieu et al. 2020 ; Isaac-Renton et al. 2018 ; Montwé et al. 2016 ). Namun, studi yang mencakup beberapa kejadian kekeringan di beberapa lokasi pengujian telah menunjukkan perbedaan yang signifikan dalam populasi yang sama dan antara lokasi dalam menanggapi kekeringan (Heer et al. 2018 ; Zas et al. 2020 ). Variabilitas dalam sifat adaptasi kekeringan di berbagai lingkungan menekankan pentingnya menganalisis respons terhadap kekeringan di berbagai lokasi untuk membuat pilihan pohon yang beradaptasi terhadap kekeringan yang kuat.
Pinus lodgepole interior ( Pinus contorta Doug. ex Loud. var. latifolia Englm.) adalah spesies konifer suksesi awal yang tersebar luas di Amerika Utara bagian barat, yang berkisar dari garis lintang 31 hingga 64° LU dan tumbuh dalam berbagai macam kondisi lingkungan (Burns 1990 ). Ini adalah salah satu spesies ekologi dan ekonomi terpenting di Kanada bagian barat, tempat beberapa lokasi uji genetik didirikan pada tahun 1970-an–1980-an. Uji genetik ini, atau kebun umum, sekarang sudah cukup tua untuk mengekstrak informasi tentang pola pertumbuhan dalam kondisi iklim yang berbeda. Dalam studi saat ini, kami menganalisis respons jangka pendek terhadap kekeringan dan pertumbuhan jangka panjang keturunan dari 40 famili penyerbukan terbuka dalam empat uji keturunan berusia 35 tahun (lokasi uji) di Kanada bagian barat dan mengeksplorasi pengaruh kondisi lingkungan dan kontrol genetik adaptasi kekeringan.
Penelitian ini mengevaluasi sifat-sifat dendrokronologis, sebagai proksi adaptasi kekeringan, dengan tiga tujuan khusus: (1) mengembangkan metrik adaptasi kekeringan yang kuat yang menggambarkan respons genotipe terhadap kekeringan; (2) menyelidiki hubungan sebab akibat di antara respons kekeringan, kerentanan penyakit, dan potensi pertumbuhan jangka panjang melalui pemodelan persamaan struktural; dan (3) menentukan tingkat kontrol genetik (varians genetik aditif) dalam sifat-sifat adaptasi kekeringan ini, dan potensinya untuk seleksi dan pemuliaan, di beberapa lokasi pengujian.
2 Bahan dan Metode
2.1 Lokasi Penelitian dan Bahan Tanaman
Sampel untuk penelitian ini berasal dari empat uji keturunan yang didirikan pada tahun 1982 (Alberta, Kanada) (Gambar 1 ). Uji coba ini awalnya ditanam untuk menguji produktivitas dan kemampuan beradaptasi berbagai famili penyerbukan terbuka (OP) dalam satu program pemuliaan. Subset famili kemudian dipilih untuk proyek seleksi genomik skala besar yang berfokus pada produktivitas dan ketahanan terhadap stres biotik dan abiotik. Di setiap lokasi, 224 famili OP (secara genetik diperlakukan sebagai famili saudara tiri karena hanya donor benih yang diketahui) dibagi menjadi 19 set, dan setiap set ditanam secara acak dalam lima blok sebagai plot baris famili empat pohon. Dengan demikian, setiap famili OP diwakili oleh 20 pohon per lokasi yang ditanam dengan jarak 2,5 × 2,5 m.
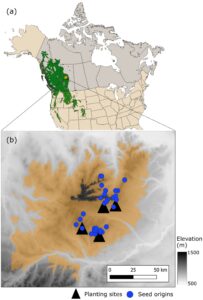
Karena lokasi penanaman dan asal pohon induk, atau provenans, semuanya berada dalam satu wilayah pembiakan, tidak ada perbedaan besar dalam kondisi iklim asli mereka, meskipun ada beberapa perbedaan elevasi kecil (Tabel 1 ). Lokasi penelitian berkisar antara 1033 hingga 1127 m di elevasi, dan rata-rata dari 2,2 ° C hingga 2,6 ° C dalam suhu tahunan rata-rata dan dari 529 hingga 555 mm dalam presipitasi selama periode penelitian (1982-2017). Kondisi iklim di asal pohon induk menunjukkan variabilitas yang lebih besar, berkisar antara 945 hingga 1280 m di elevasi, 1,2 ° C hingga 1,9 ° C dalam suhu tahunan rata-rata, dan 592 hingga 718 mm dalam presipitasi, untuk periode iklim normal 1951-1980. Jenis tanah di tiga lokasi adalah luvisolik, dengan pH 5,4 hingga 5,5, sementara lokasi keempat (lokasi Judy Creek; JUDY) memiliki tanah brunisolik yang kurang berkembang dan lebih asam dengan pH 3,9. Dua lokasi menunjukkan mortalitas yang luar biasa tinggi: VIRG (lokasi Virginia Hills) terkena kebakaran hutan pada tahun 1998 yang menewaskan sekitar setengah dari pohon yang berdiri, sementara mortalitas yang lebih tinggi dari biasanya di JUDY kemungkinan disebabkan oleh persaingan awal karena pengendalian gulma yang tidak memadai (Tabel 1 ). Semua lokasi dipagari sejak didirikan untuk mengusir ungulata, dan setiap percobaan dibatasi dengan satu baris pohon dari spesies yang sama untuk mengurangi efek tepi.
| Lokasi | Ketinggian (m) | TIKAR (°C) | (satuan ukuran) | CMI (mm) | Jenis tanah | Tingkat keasaman (pH) | Angka Kematian (%) |
|---|---|---|---|---|---|---|---|
| JUDI | tahun 1097 | 2.6 | 529 | 147 | Ortik Distrik Brunisol | 3.9 | 41.8 |
| ANGSA | tahun 1033 | 2.3 | 534 | 129 | Brunisolic abu-abu Luvisol | 5.5 | 12.6 |
| WAKTU | tahun 1064 | 2.2 | 542 | 143 | Brunisolic abu-abu Luvisol | 5.4 | 17.0 |
| PERAWAN | tahun 1127 | 2.5 | 555 | 180 | Brunisolic abu-abu Luvisol | 5.5 | 57.6 |
Singkatan: CMI, indeks kelembaban iklim; MAP, curah hujan tahunan rata-rata; MAT, suhu tahunan rata-rata.
Dari 224 famili OP yang ditanam, 40 famili, dengan sekitar 10 pohon per famili per lokasi, dipilih untuk fenotipe tingkat tinggi dan genotipe SNP ( N = 1393 pohon). Untuk mengambil sampel sejauh mana variabilitas genetik dalam penelitian, 40 famili dipilih berdasarkan nilai pemuliaan tinggi berusia 30 tahun yang mewakili famili dengan produktivitas tinggi, sedang, dan rendah. Pohon dalam famili dipilih di seluruh uji keturunan, yaitu, bukan dari plot baris tunggal famili (Cappa, Chen, et al. 2022 ; Cappa, Ratcliffe, et al. 2022 ). Bahasa Indonesia: Untuk keluarga dan pohon yang dipilih dalam keluarga, tinggi total dan diameter pada tinggi dada (DBH; 1,3 m) diukur ulang pada usia 35 (tahun 2017), dan infeksi karat empedu barat ( Endocronartium harknessii Moore) yang ada dalam percobaan dicatat pada usia 36 (tahun 2018), dan diberi skor pada skala 0 hingga 6 tergantung pada tingkat keparahan dan lokasi (cabang atau batang) empedu. Kehadiran empedu menunjukkan infeksi sebelumnya, yang mungkin telah terjadi bertahun-tahun sebelum penilaian. Indeks kompetisi dihitung menggunakan pengukuran DBH pada usia 30 (hanya 1393 pohon yang dipilih diukur pada usia 35, sementara semua 224 keluarga dan keturunannya (10.694) diukur pada usia 30), menggunakan persamaan berikut (Contreras et al. 2011 ; Rouvinen dan Kuuluvainen 1997 ):

2.2 Analisis Dendrokronologi
Pada bulan Juni 2017, satu inti utara-selatan (kulit ke kulit), dengan penambahan 5 mm diambil pada ketinggian sekitar dada (1,3 m) dari setiap pohon yang dipilih (pohon yang diinti). Profil kerapatan kayu dari bagian utara inti pohon ditentukan pada resolusi 0,0254 mm, seperti yang dijelaskan sebelumnya (Da Ros et al. 2021 ). Kami melakukan pemeriksaan kualitas yang cermat pada profil kerapatan, membuang fluktuasi kerapatan karena retakan, luka, atau ketidaksempurnaan kayu lainnya. Rata-rata kerapatan kayu dari seluruh inti dihitung dari profil kerapatan ini.
Profil kerapatan kayu dari bagian utara inti juga digunakan untuk menghitung lebar lingkaran pohon, menggunakan kontras tajam antara kerapatan kayu awal dan kayu akhir untuk mengidentifikasi lingkaran pertumbuhan tahunan. Bagian inti di mana kontras ini tidak cukup jelas untuk menentukan lebarnya dibuang (sebagian besar di lingkaran bagian dalam yang tidak tegak lurus dengan inti sehingga titik transisi yang tepat dari satu tahun ke tahun berikutnya tidak jelas) dan inti yang dibuang ini mewakili 8% (112 pohon) dari pohon yang diambil sampelnya (1393). Untuk memvalidasi pengukuran lebar lingkaran dari profil kerapatan kayu, kami menggunakan bagian selatan dari 400 inti (10 keluarga) untuk mengukur lebar lingkaran secara langsung. Inti-inti ini diampelas menggunakan butiran yang semakin halus, dipindai pada resolusi 3200 dpi (Epson Perfection V800 Photo), dan lebar lingkaran pohon diukur menggunakan WinDENDRO (versi 2017a, Regent Instruments Canada Inc.). Semua pengukuran lebar lingkaran pertama-tama diperiksa tanggalnya secara visual dan kemudian diperiksa kesalahannya menggunakan COFECHA (Holmes 1983 ). Nilai lebar lingkaran kemudian digunakan untuk menghitung kenaikan luas basal tahunan (BAI) menggunakan fungsi bai.out dalam paket dplR untuk R (Bunn 2008 ). Hasil pengukuran yang berasal dari satu dan dua inti per pohon dibandingkan dan perbandingan ini tidak menghasilkan perbedaan yang signifikan, jadi kami berasumsi bahwa satu inti secara akurat mewakili pertumbuhan pohon.
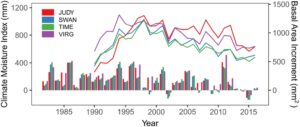
Berdasarkan data pertumbuhan dan iklim, kami memilih dua kejadian kekeringan yang berbeda, di mana pohon menunjukkan penurunan pertumbuhan yang jelas, untuk menghitung beberapa indeks respons kekeringan (Gambar 2 ). Kejadian kekeringan ini dicirikan oleh akhir musim tanam yang sangat kering pada tahun sebelumnya (Agustus/September 2001 dan 2009), diikuti oleh musim panas yang sangat kering (2002) dan awal musim tanam yang kering (April–Juni, 2010; Gambar S1 ), masing-masing. Meskipun periode kekeringan lain terjadi pada 2014/2015, itu tidak termasuk dalam analisis ini karena, pada saat pengumpulan sampel pada 2017, tidak ada cukup tahun pascakekeringan untuk mengukur pemulihan pertumbuhan. Kami menghitung indeks resistensi dan ketahanan untuk setiap kejadian kekeringan sebagai berikut (Lloret et al. 2011 ):

di mana, kejadian BAI = BAI tahun kejadian kekeringan, BAI pra = BAI rata-rata empat tahun sebelum kejadian kekeringan, dan BAI pasca = BAI rata-rata empat tahun setelah kejadian kekeringan. Kami memilih rata-rata empat tahun sebagai kompromi antara memiliki sebanyak mungkin tahun untuk mendapatkan representasi yang baik dari pertumbuhan rata-rata sambil menghindari kejadian kekeringan kecil lainnya.
Karena ketahanan dan resistensi merupakan respons jangka pendek pohon terhadap kekeringan, kami menggunakan pendekatan serupa untuk mengembangkan indeks sederhana baru yang akan mewakili respons jangka panjang pohon terhadap kekeringan berulang. Karena pohon yang diteliti dalam uji coba ini memiliki jarak yang sama dan usia yang sama (yaitu, mengikuti pola pertumbuhan yang sama), kami membandingkan pertumbuhan selama periode lima tahun produktivitas maksimum, tepat sebelum kekeringan parah pertama (1997–2001), dan pertumbuhan selama lima tahun terakhir yang tercatat (2012–2016), sebagai berikut:
![]()
di mana, BAI maks = BAI rata-rata selama 1997–2001, dan BAI terakhir = BAI rata-rata antara 2012–2016. Pohon yang menunjukkan nilai penurunan mendekati 1 mewakili individu yang tidak mengalami penurunan yang mampu mempertahankan pola pertumbuhan yang konstan sepanjang masa hidupnya, sementara pohon dengan nilai penurunan yang lebih besar (> 1) mewakili individu yang mengalami penurunan pertumbuhan yang parah dibandingkan dengan pertumbuhan awal mereka. Kami memilih rata-rata lima tahun untuk indeks ini dengan mengikuti alasan yang sama yang dijelaskan di atas, meskipun 2015, tahun kekeringan, dimasukkan dalam BAI terakhir .
2.3 Analisis δ 13 C
Kami menggunakan dua lempengan sisa kecil, yang diambil dari panjang penuh bagian utara setiap inti 5 mm, untuk melakukan analisis δ 13 C. Sampel inti dikeringkan pada suhu 65°C selama 3 hari dan digiling menggunakan Qiagen Tissuelyser II (Qiagen Inc.-Canada, Ontario, Kanada). Sampel kayu giling selanjutnya dikirim ke Stable Isotope Lab milik InnoTech Alberta (Victoria, BC, Kanada) untuk analisis δ 13 C, tanpa ekstraksi selulosa (Isaac-Renton et al. 2018 ; Hu et al. 2021 ). Sampel dianalisis menggunakan metode yang sudah mapan pada spektrometer massa MAT253 dengan antarmuka Conflo IV (Thermo Fisher Scientific, Waltham, MA, AS) dan Fisons NA1500 EA (Fisons Instruments, Milano, Italia). Hasilnya dinormalisasi dan dilaporkan terhadap Vienna Pee Dee Belemnite (standar yang ditetapkan untuk estimasi karbon-13 [δ 13 C] [Craig 1957 ]).
2.4 Iklim
Kami menggunakan variabel bulanan dan tahunan dari tahun 1982 hingga 2016 untuk menggambarkan kondisi iklim yang dialami pohon sejak didirikan. Untuk mengkarakterisasi iklim asal pohon induk, kami mengekstrak rata-rata untuk periode normal 1950–1980 menggunakan perangkat lunak BioSIM v.11.3 (Régnière et al. 2017 ), yang merupakan periode normal terbaru sebelum uji coba didirikan. Data kemudian diinterpolasi dari empat stasiun cuaca terdekat yang tersedia dan dikoreksi untuk lokasi geografis dan ketinggian. Variabel yang digunakan dalam penelitian ini adalah suhu tahunan rata-rata (MAT), curah hujan tahunan rata-rata (MAP), dan indeks kelembapan iklim (CMI = curah hujan – evapotranspirasi Hargraves).
2.5 Rekonstruksi dan Koreksi Silsilah
Karena sifat koleksi benih penyerbukan terbuka, verifikasi silsilah diperlukan menggunakan penanda SNP yang tersedia (Cappa, Chen, et al. 2022 ). Kesalahan silsilah diidentifikasi dan dikoreksi menggunakan skrip R khusus untuk membandingkan estimasi hubungan aditif berpasangan yang diamati dengan nilai yang diharapkan (misalnya, 0,25 untuk saudara tiri). Estimasi hubungan aditif berpasangan yang diamati dihitung menurut VanRaden ( 2008 ). Keputusan untuk menetapkan sampel ke kelompok genetik (keluarga silsilah) didasarkan pada hubungan rata-rata maksimum yang diamati dari sampel dengan setiap kelompok genetik (lihat Gambar S2 dalam Cappa, Ratcliffe, et al. 2022 ).
2.6 Analisis Statistik
Nilai rata-rata dendrokronologi dan sifat pertumbuhan untuk setiap lokasi dihitung menggunakan model campuran di mana lokasi dianggap sebagai efek tetap dan blok dan set yang bersarang di dalam blok dianggap sebagai efek acak. Jika memungkinkan, hanya satu pohon yang diambil sampelnya dari setiap plot empat baris pohon; oleh karena itu, efek plot tidak disesuaikan. Model-model ini digunakan untuk menghitung estimasi rata-rata marginal untuk setiap sifat, dan perbedaan signifikan antara lokasi dianalisis menggunakan perbandingan berpasangan dengan koreksi Tukey. Hubungan fenotipik linear antara sifat dianalisis dengan model linear dan korelasi momen-produk Pearson, dan nilai- p korelasi dikoreksi untuk beberapa perbandingan menggunakan penyesuaian Bonferroni. Dalam semua kasus, kenormalan data diperiksa, apriori , dan transformasi akar kuadrat atau logaritmik diterapkan jika diperlukan.
Hubungan kompleks antara sifat-sifat dianalisis lebih lanjut menggunakan model persamaan struktural (SEM). SEM ini adalah analisis konfirmatori yang menguji apakah model teoritis yang didefinisikan melalui hubungan kausal langsung dan tidak langsung antara sifat-sifat dapat dengan tepat menjelaskan varians dalam data, sambil mengidentifikasi jika ada hubungan penting yang hilang (Lefcheck 2016 ). Dalam kasus ini, kami menggunakan SEM asiklik terarah (atau sepotong-sepotong), yang mengacu pada aplikasi dari teori grafik. Pendekatan ini memungkinkan kami untuk bekerja dengan data yang tidak normal dan tidak independen dan menghitung hubungan bivariat dalam model menggunakan model campuran linier umum. Oleh karena itu, hubungan dalam model dikoreksi untuk efek acak dari desain eksperimen (situs, blok, dan set) dan genetika (keluarga). Kebaikan kecocokan SEM dihitung menggunakan statistik Fisher C yang mengikuti distribusi chi-kuadrat, dan nilai – p di atas 0,05 menunjukkan bahwa model tersebut tidak kehilangan hubungan yang signifikan. Keuntungan lain dari metode SEM adalah menguji pengaruh semua tautan yang tidak didefinisikan dalam model asli, sehingga mengidentifikasi hubungan signifikan yang awalnya tidak diharapkan. Kekuatan hubungan linier individual dalam model diwakili oleh koefisien standar (SC) yang berkisar antara -1 dan +1. SEM dihitung menggunakan paket piecewiseSEM untuk R (Lefcheck 2016 ).
Estimasi heritabilitas arti sempit dan korelasi genetik antara sifat dan lokasi dihitung dari model campuran individu-pohon multi-sifat dengan persamaan berikut menggunakan algoritma Ekspektasi–Maksimalisasi, diikuti oleh satu putaran dengan algoritma Informasi-Rata, untuk menghitung kesalahan standarnya dalam paket breedR untuk R (Muñoz dan Sanchez 2019 ):
![]()

3 Hasil
3.1 Respon Kekeringan di Berbagai Lokasi Uji
Peristiwa kekeringan pada tahun 2001–2002 dan 2009–2010 menunjukkan dampak negatif pada pertumbuhan pohon di keempat lokasi uji keturunan (Gambar 2 ). Secara keseluruhan, peristiwa kekeringan tahun 2002 tampak lebih signifikan daripada peristiwa kekeringan tahun 2010 karena nilai ketahanan dan resistansi secara konsisten lebih rendah pada tahun 2002, kecuali untuk ketahanan di lokasi JUDY (Tabel 2 ). Selain itu, ada perbedaan signifikan antara lokasi dalam respons mereka terhadap kekeringan. Lokasi Swan Hills (SWAN) dan Timeau (TIME) menunjukkan pengurangan pertumbuhan yang lebih kuat karena kekeringan, menunjukkan penurunan yang umumnya lebih tinggi dan nilai resistansi dan ketahanan yang lebih rendah, dibandingkan dengan lokasi JUDY dan VIRG (Tabel 2 ). Perbedaan-perbedaan ini sangat kuat dalam ketahanan terhadap peristiwa kekeringan tahun 2002 dan dalam penurunan jangka panjang. Situs SWAN dan TIME menunjukkan ketahanan terendah pada tahun 2002 (rata-rata 0,78 dan 0,75 di setiap situs, berturut-turut) dan penurunan tertinggi (rata-rata 1,83 untuk SWAN dan TIME), diikuti oleh VIRG (penurunan rata-rata 1,52 dan ketahanan rata-rata 0,82 pada tahun 2002), dan kemudian JUDY, yang paling sedikit terpengaruh (penurunan 1,38 dan ketahanan 0,91 pada tahun 2002). Kami mengamati pola serupa dalam pertumbuhan di JUDY, yang menunjukkan tinggi tertinggi dan DBH tertinggi kedua (masing-masing 1371 cm dan 19,4 cm). VIRG menunjukkan tinggi tertinggi kedua dan DBH yang lebih besar (1332 cm dan 20,1 cm) dan SWAN dan TIME menunjukkan nilai terendah pada kedua sifat (1249 cm dan 18,3 cm pada SWAN, dan 1268 dan 17,6 pada TIME, berturut-turut) (Tabel 2 ).
| Sifat | JUDI | ANGSA | WAKTU | PERAWAN |
|---|---|---|---|---|
| Menolak | 1,38 (0,03) suatu | 1,83 (0,04) detik | 1,83 (0,04) detik | 1,52 (0,04)b |
| Ketahanan 2002 | 0,91 (0,01) suatu | 0,78 (0,01) SM | 0,75 (0,01) detik | 0,82 (0,01)b |
| Ketahanan 2010 | 0,87 (0,01)b | 0,78 (0,01) detik | 0,86 (0,01)b | 0,92 (0,02) suatu |
| Perlawanan 2002 | 0,75 (0,01) suatu | 0,74 (0,01) persen | 0,71 (0,01) detik | 0,72 (0,01) SM |
| Perlawanan 2010 | 0,83 (0,01) suatu | 0,74 (0,01)b | 0,87 (0,01) suatu | 0,87 (0,01) suatu |
| Tinggi 35 (cm) | 1371 (8.5) sebuah | 1249 (7.8) hari | 1268 (7.9) hari | 1332 (9.2) tahun |
| Tinggi Badan 35 (cm) | 19,4 (0,16)b | 18.3 (1.15) detik | 17,6 (0,15) hari | 20.1 (0.18) sebuah |
Kami menguji apakah pertumbuhan sebelum peristiwa kekeringan memiliki efek pada respons pohon terhadap kekeringan dan menemukan hubungan positif minor antara ketahanan terhadap peristiwa kekeringan 2002 dan pertumbuhan sebelumnya ( p = 0,002), yang lebih kuat di lokasi SWAN ( p = 0,008) dan TIME ( p = 0,015) (Gambar S2 ). Hasil-hasil ini juga menunjukkan efek negatif signifikan dari pertumbuhan sebelumnya dalam ketahanan terhadap peristiwa kekeringan 2010 ( p = 0,044), yang dalam kasus ini lebih kuat di JUDY ( p = 0,059) dan TIME ( p = 0,080). Tiga indeks respons kekeringan lainnya hanya menunjukkan hubungan signifikan dengan pertumbuhan sebelumnya di salah satu lokasi, tetapi dalam semua kasus, pohon dengan pertumbuhan yang lebih tinggi sebelum peristiwa kekeringan lebih parah dipengaruhi oleh stres kekeringan. Di JUDY, pohon dengan pertumbuhan yang lebih tinggi sebelum tahun 2002 memiliki ketahanan yang lebih rendah terhadap peristiwa kekeringan 2002 dan penurunan jangka panjang yang lebih tinggi. Pada TIME, pohon dengan pertumbuhan lebih besar sebelum peristiwa kekeringan 2010 juga menunjukkan ketahanan yang lebih rendah terhadap kekeringan. Pertumbuhan sebelum 2002 juga tidak menunjukkan efek pada ketahanan berikutnya terhadap beberapa peristiwa kekeringan (Gambar 3 ). Namun, kemampuan untuk pulih, atau tidak, setelah satu atau dua peristiwa stres kekeringan memiliki efek yang kuat pada pertumbuhan berikutnya dari pohon-pohon yang dipelajari di keempat lokasi (Gambar 3 , Gambar S3 ). Pohon dengan ketahanan tinggi terhadap kedua peristiwa kekeringan mampu mempertahankan laju pertumbuhannya sebelum dan setelah setiap peristiwa kekeringan (Gambar 3b,c ).
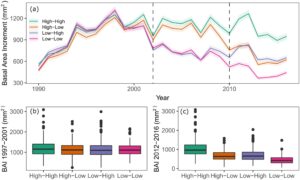
3.2 Respon terhadap Kekeringan dan Pertumbuhan Jangka Panjang
Kami menganalisis korelasi fenotipik (Gambar 4a ) dan genetik (Gambar 4b ) antara indeks respons kekeringan dan kinerja jangka panjang. Kemampuan pohon untuk pulih dari peristiwa kekeringan, mempertahankan laju pertumbuhan yang mirip dengan yang tercatat sebelum peristiwa tersebut, memiliki efek yang kuat pada pertumbuhan jangka panjang pohon (Gambar 4a ). Sementara pohon dengan ketahanan yang lebih tinggi terhadap peristiwa kekeringan tahun 2002 dan 2010 memiliki penurunan jangka panjang yang jauh lebih rendah ( masing-masing R = −0,60, p < 0,001 dan R = −0,57, p < 0,001). Kemampuan untuk mempertahankan pertumbuhan selama peristiwa kekeringan memiliki dampak yang lebih rendah, tetapi signifikan, pada keberhasilan jangka panjang pohon dengan nilai korelasi sebesar -0,29 ( p < 0,001) dan -0,31 ( p < 0,001) antara penurunan dan ketahanan terhadap peristiwa kekeringan tahun 2002 dan 2010, masing-masing. Dua indeks respons jangka pendek, ketahanan dan ketangguhan, sangat berkorelasi untuk peristiwa kekeringan yang sama ( R = 0,65, p < 0,001 pada tahun 2002 dan R = 0,60, p < 0,001 pada tahun 2010). Sebaliknya, ketahanan ( R = -0,09, p = 0,003) dan ketangguhan ( R = -0,13, p < 0,001), yang sesuai dengan berbagai peristiwa kekeringan, berkorelasi negatif (Gambar 4a ).

Pohon yang menunjukkan respons kekeringan yang lebih baik menunjukkan pertumbuhan akhir yang lebih tinggi (Gambar 4a ). Penurunan jangka panjang menunjukkan korelasi tertinggi dengan tinggi dan DBH pada usia 35 ( R = −0,33, p < 0,001, dan R = −0,36, p < 0,001, berturut-turut). Di antara dua indeks jangka pendek, ketahanan, dengan nilai korelasi dari 0,13 hingga 0,31, memiliki efek yang lebih kuat daripada resistensi (0,03–0,21) pada pertumbuhan akhir (tinggi dan DBH) pohon setelah kedua peristiwa kekeringan (Gambar 4a ). Untuk dua peristiwa kekeringan, kekeringan 2002 menunjukkan efek yang lebih kuat pada pertumbuhan akhir (0,08–0,31) daripada peristiwa kekeringan 2010 (0,03–0,18, Gambar 4 ) baik di seluruh ketahanan maupun resistensi. Selanjutnya, semua korelasi fenotipik mengikuti tren yang sangat mirip di keempat lokasi (Gambar S4 ).
Kami selanjutnya menjelaskan hubungan antara indeks respons kekeringan, pertumbuhan, fisiologis, dan variabel lingkungan dengan SEM sepotong-sepotong dengan Fisher’s C sebesar 32,22 dan p = 0,555 (Gambar 5 ). Beberapa tren yang diidentifikasi di bagian sebelumnya juga hadir dalam SEM. Pohon yang kurang tahan kekeringan terhadap salah satu dari dua peristiwa kekeringan mengalami penurunan jangka panjang yang lebih tinggi yang, pada gilirannya, mengurangi pertumbuhan akhir pohon seperti yang diwakili oleh tinggi dan DBH, yang merupakan sifat berkorelasi (Gambar 5b ). Meskipun ketahanan terhadap peristiwa kekeringan 2002 dan 2010 adalah variabel dengan pengaruh terbesar pada penurunan, dengan koefisien standar (SC) masing-masing -0,65 dan -0,66, tekanan lain berkontribusi, dengan intensitas yang lebih rendah, terhadap penurunan keseluruhan pohon. Pohon yang menderita persaingan lebih banyak mengalami penurunan yang lebih kuat dengan SC 0,10, dan pohon yang menderita infeksi karat empedu barat yang lebih parah juga menunjukkan penurunan yang lebih tinggi (SC = 0,04). Pohon yang menderita persaingan lebih banyak menunjukkan ketahanan yang lebih rendah terhadap kedua peristiwa kekeringan (SC = -0,26 dan SC = -0,12 masing-masing pada tahun 2002 dan 2010), dan pohon dengan kehadiran karat empedu barat yang lebih besar tampaknya kurang tahan terhadap kekeringan pada tahun 2002 (SC = -0,24). Pohon yang ditekan juga menunjukkan kepadatan kayu yang lebih tinggi (SC = 0,23). Kami berhipotesis, dalam model teoritis kami, bahwa δ 13 C, sebagai proksi untuk efisiensi penggunaan air, dan kepadatan kayu, sebagai proksi untuk resistensi kavitasi, akan mendukung ketahanan kekeringan, tetapi tidak satu pun dari hubungan ini ditemukan signifikan dalam SEM.
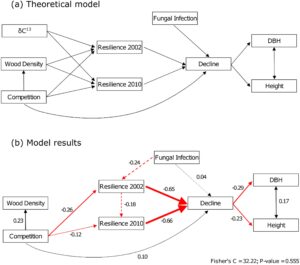
3.3 Heritabilitas dan Kontrol Genetik Sifat Adaptif terhadap Kekeringan
Kelima indeks respon kekeringan menunjukkan estimasi heritabilitas rendah hingga sedang (
) di setiap lokasi, sementara juga menunjukkan estimasi yang relatif rendah ketika keempat lokasi digabungkan, yang menunjukkan adanya interaksi G × E (Tabel 3 ). Penurunan menunjukkan estimasi tertinggi untuk lima indeks respons kekeringan berkisar antara 0,15 hingga 0,30, di seluruh lokasi. Di antara sifat jangka pendek, ketahanan tampaknya merupakan sifat yang lebih dapat diwariskan daripada ketahanan, meskipun ketahanan VIRG 2010 merupakan estimasi heritabilitas tunggal tertinggi (
= 0,39 ± 0,06) (Tabel 3 ). Sifat pertumbuhan menunjukkan estimasi heritabilitas yang lebih tinggi dibandingkan sifat respon kekeringan, baik pada tingkat lokasi individu maupun lokasi gabungan, dengan tinggi badan menjadi sifat yang paling dapat diwariskan (
= 0,40–0,81) diikuti oleh DBH (
= 0,20–0,64) (Tabel 3 ). Estimasi yang lebih rendah pada data gabungan situs disebabkan oleh G × E, sebagaimana dibuktikan oleh korelasi genetik yang buruk antara situs (Tabel S1 ). Sebagian besar sifat tidak berkorelasi secara signifikan antara situs (Tabel S1 ). Namun, SWAN dan TIME, situs dengan kondisi lingkungan yang paling mirip (Tabel 1 ), menunjukkan korelasi tertinggi dalam kemunduran (0,81), ketahanan 2002 (0,73), dan resistensi 2002 (0,78). Satu-satunya korelasi signifikan lainnya adalah antara JUDY dan SWAN untuk ketahanan 2010 (0,73).
| Sifat | Semua situs | JUDI | ANGSA | WAKTU | PERAWAN |
|---|---|---|---|---|---|
| Menolak | 0,12 (0,03) | 0,30 (0,12) | 0,24 (0,04) | 0,30 (0,08) | 0,15 (0,02) |
| Ketahanan 2002 | 0,09 (0,03) | 0,27 (0,11) | 0,29 (0,04) | 0,15 (0,06) | 0,07 (0,01) |
| Ketahanan 2010 | 0,06 (0,02) | 0,08 (0,05) | 0,25 (0,03) | 0,21 (0,06) | 0,25 (0,04) |
| Perlawanan 2002 | 0,06 (0,02) | 0,19 (0,08) | 0,16 (0,02) | 0,17 (0,07) | 0,03 (0,01) |
| Perlawanan 2010 | 0,03 (0,01) | 0,12 (0,08) | 0,03 (0,01) | 0,11 (0,05) | 0,39 (0,06) |
| DBH 35 | 0,29 (0,05) | 0,47 (0,14) | 0,22 (0,03) | 0,20 (0,06) | 0,64 (0,08) |
| Tinggi 35 | 0,50 (0,06) | 0,54 (0,15) | 0,81 (0,08) | 0,50 (0,14) | 0,40 (0,06) |
Korelasi genetik yang signifikan antara sifat-sifat mengikuti tren yang sama seperti korelasi fenotipik dengan analisis gabungan situs (Gambar 4 ) dan analisis situs individual (Gambar S4 dan S5 ). Ketahanan, terhadap kejadian tahun 2002 dan 2010, berkorelasi signifikan dengan penurunan jangka panjang ( masing-masing R = −0,78 dan −0,82; Gambar 4b ). Penurunan tersebut juga berkorelasi kuat dengan kedua sifat pertumbuhan ( tinggi R = −0,62 dan DBH R = −0,55). Lebih jauh lagi, tinggi berkorelasi signifikan dengan ketahanan 2010 ( R = 0,57) dan DBH berkorelasi dengan ketahanan 2002 ( R = 0,60) (Gambar 4b ).
Akhirnya, kami mengamati adaptasi terhadap lingkungan lokal dalam ketahanan terhadap peristiwa kekeringan tahun 2002 dan penurunan jangka panjang. Kedua sifat tersebut berkorelasi secara signifikan dengan ketinggian asal (provenance) pohon induk donor ( R2 = 0,20, p = 0,002 untuk ketahanan tahun 2002 dan R2 = 0,16, p = 0,007 untuk penurunan; Gambar 6 ). Pohon yang berasal dari ketinggian yang lebih rendah dengan suhu yang lebih tinggi dan curah hujan yang lebih rendah (yaitu, kondisi yang lebih kering) menunjukkan peningkatan respons kekeringan.

4 Diskusi
Studi ini menganalisis ketahanan dan resistensi terhadap kekeringan dari 40 famili OP pinus lodgepole berusia 35 tahun yang tumbuh di empat lokasi uji keturunan yang mengalami beberapa peristiwa kekeringan. Di bawah peristiwa kekeringan yang sering terjadi pada paruh kedua periode pertumbuhan, pohon yang menunjukkan ketahanan kekeringan jangka pendek yang tinggi (hingga empat tahun setelah peristiwa kekeringan) menunjukkan penurunan pertumbuhan jangka panjang yang lebih rendah, dan mencapai tinggi akhir dan diameter setinggi dada (DBH) yang lebih unggul. Hubungan positif antara ketahanan terhadap kekeringan dan pertumbuhan akhir ini bertentangan dengan pengamatan yang lebih umum yang ditemukan di mana terdapat trade-off antara respons kekeringan dan ukuran pohon (Bennett et al. 2015 ; Merlin et al. 2015 ; Serra-Maluquer et al. 2018 ). Studi sebelumnya menunjukkan bahwa pohon yang lebih besar biasanya lebih rentan terhadap stres kekeringan, dan ini dikaitkan dengan alokasi karbon yang lebih tinggi untuk biomassa di atas tanah, kerentanan hidrolik yang lebih tinggi, permintaan evaporatif yang lebih tinggi karena status sosial tegakannya, dan preferensi kumbang kulit kayu untuk menyerang pohon yang lebih besar (Serra-Maluquer et al. 2018 ). Namun, temuan yang bertentangan juga telah dilaporkan, yang menunjukkan bahwa pohon yang lebih besar mungkin memiliki sistem akar yang substansial, yang mendukung perolehan air selama kekeringan (Hember et al. 2017 ; Zang et al. 2012 ). Studi yang disebutkan di atas mempertimbangkan respons kekeringan terhadap ukuran pohon. Dalam studi ini, kami telah menunjukkan bahwa pohon memiliki kemampuan untuk pulih dari satu atau beberapa peristiwa kekeringan pada tahap awal kehidupan, yang pada gilirannya dapat memengaruhi pertumbuhan jangka panjang pohon. Faktanya, kami menemukan sedikit atau tidak ada hubungan antara respons terhadap satu atau beberapa peristiwa kekeringan dan pertumbuhan pohon sebelum peristiwa kekeringan (Gambar 3 , Gambar S2 ). Hasil penelitian kami menunjukkan bagaimana pohon yang tidak tahan kekeringan dapat mengalami pengurangan pertumbuhan jangka panjang yang parah jika kejadian kekeringan terus meningkat dalam frekuensi dan tingkat keparahan, yang menyebabkan peningkatan kematian pohon, dan akhirnya berdampak pada siklus karbon global dalam menghadapi perubahan iklim (Anderegg et al. 2015 ).
Jika adaptasi kekeringan sebagai prioritas dalam pengelolaan hutan diinginkan, pengetahuan tentang mekanisme yang mendasari adaptasi kekeringan baik dalam jangka pendek maupun jangka panjang sangat penting. Hasil kami menunjukkan dampak yang lebih besar dari ketahanan terhadap pertumbuhan dan penurunan jangka panjang daripada resistensi (Gambar 4 ). Prevalensi ketahanan ini sebagai strategi adaptasi kekeringan sebelumnya telah dilaporkan pada provenans pinus maritim ( Pinus pinaster Ait.) yang beradaptasi dengan kondisi yang lebih kering (Sánchez-Salguero et al. 2018 ). Ketahanan yang lebih tinggi telah terbukti berhubungan dengan mortalitas yang lebih rendah akibat kekeringan pada konifer, seperti yang disorot dalam studi global, yang kontras dengan pengaruh yang lebih besar dari resistensi kekeringan pada angiospermae (DeSoto et al. 2020 ). Dalam skenario perubahan iklim, yang dicirikan oleh peristiwa kekeringan ringan hingga parah yang berulang, seperti iklim yang dialami dalam studi ini, kemampuan untuk pemulihan yang cepat setelah stres kekeringan menjadi penting dalam mencegah penurunan pertumbuhan jangka panjang. Hipotesis alternatif adalah bahwa ketahanan yang lebih rendah, seperti yang diukur dengan pemulihan ke tingkat pertumbuhan sebelum kekeringan, dapat mengindikasikan adaptasi plastik terhadap kekeringan. Plastisitas ini dapat menyebabkan pohon mengorbankan pertumbuhan di atas tanah sebagai ganti pengembangan jaringan yang tahan kekeringan dan sistem akar yang lebih besar (Corcuera et al. 2004 ; Eilmann et al. 2009 ; Gessler et al. 2020 ; Hagedorn et al. 2016 ).
Lebih jauh lagi, studi kami mengungkap korelasi negatif antara resistensi dan ketahanan pada tingkat fenotipik dalam dua peristiwa kekeringan yang dipelajari, memberikan dukungan pada hipotesis bahwa pemulihan yang lambat dari satu peristiwa kekeringan dapat menyebabkan peningkatan plastisitas dalam respons kekeringan terhadap peristiwa kekeringan berikutnya (Reed et al. 2011 ). Namun, perlu juga dicatat interpretasi alternatif dari hasil ini. Pertama, korelasi genetik negatif yang kuat antara ketahanan terhadap peristiwa kekeringan 2002 dan penurunan jangka panjang ( R = −0,78; p < 0,001, Gambar 4b ) menunjukkan bahwa 14 tahun setelah peristiwa kekeringan, pohon masih berjuang untuk pulih ke tingkat pertumbuhan sebelum kekeringan. Studi lain tentang mortalitas akibat kekeringan telah menunjukkan tren penurunan pertumbuhan yang serupa selama lebih dari 20 tahun sebelum kematian pohon, yang menekankan pentingnya mempertahankan tingkat pertumbuhan yang konsisten untuk bertahan hidup dari peristiwa kekeringan di masa mendatang (Camarero et al. 2015 ; DeSoto et al. 2020 ). Pertimbangan kedua adalah bahwa korelasi genetik yang cukup positif antara ketahanan terhadap peristiwa kekeringan 2002 dan 2010 ( R = 0,40; SE = 0,22, Gambar 4b ), meskipun tidak signifikan secara statistik, menunjukkan bahwa pohon yang beradaptasi dengan kekeringan umumnya menunjukkan ketahanan yang lebih baik terhadap beberapa peristiwa kekeringan. Korelasi fenotipik negatif yang diamati dapat dijelaskan oleh kasus ekstrem di mana pohon yang mengimbangi pertumbuhannya secara berlebihan setelah kekeringan 2002 (dengan nilai ketahanan lebih tinggi dari 1) sangat terpengaruh oleh peristiwa 2010, atau pohon yang tidak pulih setelah 2002 dan mempertahankan tingkat pertumbuhan yang rendah (yaitu, ketahanan tinggi) setelah peristiwa kekeringan 2010.
Upaya penelitian yang cukup besar telah diinvestasikan dalam menemukan sifat adaptif yang terkait dengan respons kekeringan, termasuk asosiasi dengan sifat seperti kepadatan kayu yang tinggi dan luas daun spesifik (Greenwood et al. 2017 ), atau plastisitas yang lebih tinggi dalam anatomi xilem dan efisiensi penggunaan air (Isaac-Renton et al. 2018 ). Lebih jauh lagi, kepadatan kayu yang lebih tinggi, misalnya, sering dikaitkan dengan trakeid dengan area lumen yang lebih kecil dan dinding sel yang lebih tebal, memberikan ukuran ketahanan terhadap emboli di bawah tekanan kekeringan (Hacke et al. 2001 ; Sperry et al. 2006 ). Sementara kami awalnya berhipotesis bahwa pohon pinus lodgepole dengan kepadatan kayu yang lebih tinggi akan menunjukkan ketahanan dan ketahanan kekeringan yang lebih besar, hasil kami tidak mendukung harapan ini (Gambar 5 ), yang selanjutnya mengonfirmasi hasil yang diperoleh oleh Da Ros et al. ( 2021 ), menggunakan set data yang sama. Hubungan lemah antara kerapatan kayu dan ketahanan dan ketahanan kekeringan juga telah terlihat pada spesies Abies (George et al. 2015 ), yang menunjukkan bahwa sifat xilem mungkin bukan sifat yang paling relevan untuk menilai adaptasi kekeringan pada konifer. Choat et al. ( 2012 ) menyatakan bahwa sebagian besar spesies pohon hutan beroperasi pada margin keamanan hidrolik yang rendah (yaitu, perbedaan antara potensial air umum dalam xilem dan potensial air minimum yang dapat menyebabkan kerusakan xilem), terlepas dari sifat xilem atau kondisi iklim. Ini menyiratkan bahwa kemampuan untuk mengatur penutupan stomata dan meningkatkan efisiensi penggunaan air selama peristiwa kekeringan dapat menjadi kunci pada pinus lodgepole. Isaac-Renton et al. ( 2018 ) menemukan bahwa asal usul pinus lodgepole yang beradaptasi dengan kekeringan dapat menyesuaikan perilaku stomata dalam kondisi kekeringan, yang memungkinkan asal usul ini mencapai pertumbuhan akhir yang lebih tinggi. Meskipun kami tidak mengidentifikasi korelasi antara respons pohon terhadap kekeringan dan efisiensi penggunaan air, sebagaimana diukur dengan δ 13 C, data ini diperoleh dari rata-rata keseluruhan periode pertumbuhan yang ditangkap oleh inti kayu yang diambil setinggi dada (sekitar usia 20–35), sehingga menjadi tantangan untuk mengetahui potensi respons plastik terhadap peristiwa kekeringan tertentu.
Faktor ekstrinsik, di luar peristiwa kekeringan itu sendiri, juga ditemukan memengaruhi kemampuan pohon untuk melawan dan pulih dari tekanan kekeringan. Studi kami mengamati bahwa persaingan dan kerentanan terhadap karat empedu barat mengurangi ketahanan kekeringan dan meningkatkan nilai penurunan jangka panjang (Gambar 5 ). Sementara pohon yang lebih kecil mungkin mengalami lebih sedikit tekanan kekeringan karena tempat berlindung yang disediakan oleh pohon yang lebih dominan (Aussenac 2000 ), kalah bersaing untuk mendapatkan sumber daya dapat menghambat kemampuan mereka untuk pulih setelah peristiwa kekeringan. Persaingan dapat memiliki dampak yang sebanding atau bahkan lebih besar daripada iklim pada tren pertumbuhan jangka panjang (Gleason et al. 2017 ; Young et al. 2017 ; Zhang et al. 2015 ), namun, itu juga dapat memiliki efek yang kuat pada pemulihan jangka pendek setelah kekeringan. Dalam studi kami, dua lokasi dengan ~50% mortalitas sebelum peristiwa kekeringan pertama, dan oleh karena itu persaingan yang lebih rendah, menunjukkan ketahanan yang jauh lebih tinggi dan penurunan yang lebih rendah. Pengamatan ini menunjukkan bahwa pengelolaan jarak yang lebih luas (tegalan lebih terbuka), dengan persaingan yang berkurang, bisa menjadi strategi untuk mengurangi dampak kekeringan pada pertumbuhan pohon, sehingga menghasilkan hutan yang tahan kekeringan (D’Amato et al. 2013 ; Sohn et al. 2013 , 2016 ).
Stres abiotik (misalnya, kekeringan) dan biotik (misalnya, penyakit) dapat berinteraksi secara sinergis, berfungsi untuk mempengaruhi dan memicu faktor-faktor, yang mengakibatkan penurunan atau kematian jangka panjang (McDowell et al. 2008 ). Hasil kami menunjukkan bahwa pohon yang terinfeksi parah oleh karat empedu barat menunjukkan respons kekeringan yang berkurang. Dampak negatif karat empedu barat pada sifat hidrolik pinus lodgepole dipelajari oleh Wolken et al. ( 2009 ), yang menunjukkan bahwa, dalam kondisi basah, pohon yang terkena dampak memiliki kapasitas hidrolik yang lebih rendah sementara pohon yang terinfeksi dan sehat sama-sama terkena dampak kekeringan. Pengamatan ini selanjutnya menjelaskan mengapa pohon yang terkena karat empedu barat tampak terganggu dalam kemampuannya untuk pulih ke tingkat pertumbuhan sebelum kekeringan dibandingkan dengan pohon yang sehat, karena mereka mungkin tidak dapat memanfaatkan sepenuhnya periode dengan ketersediaan air yang memadai.
Beberapa studi konifer baru-baru ini telah menganalisis adaptasi genetik terhadap kekeringan menggunakan sifat-sifat dendrokronologis. Beberapa studi menunjukkan hubungan antara sifat-sifat dendrokronologis dan penanda molekuler fungsional (Heer et al. 2018 ; Housset et al. 2018 ; Trujillo-Moya et al. 2018 ), sementara yang lain melaporkan adaptasi yang kuat terhadap kondisi lokal ketika sumber benih berasal dari daerah yang lebih kering (Depardieu et al. 2020 ; Montwé et al. 2016 ). Studi-studi ini menyoroti potensi penggunaan sifat-sifat berbasis lingkaran pohon, seperti ketahanan, dalam seleksi genom. Di sini, kami menemukan estimasi heritabilitas rendah hingga sedang dalam model genetik satu lokasi, dengan heritabilitas yang relatif lebih rendah dari semua lokasi yang digabungkan (Tabel 3 ). Dalam semua kasus, estimasi heritabilitas dari lima sifat respons kekeringan lebih rendah daripada sifat-sifat pertumbuhan klasik, seperti tinggi dan DBH. Nilai heritabilitas serupa dalam indeks jangka pendek di satu lokasi juga dilaporkan dalam studi cemara putih ( Picea glauca (Moench) Voss) baru-baru ini (Depardieu et al. 2020 ). Estimasi heritabilitas yang lebih rendah dalam model lokasi gabungan dijelaskan oleh korelasi yang rendah antara lokasi (yaitu, G × E). Indeks berbasis lingkaran pohon mencerminkan reaksi pohon terhadap kondisi lingkungan tertentu, yang berkontribusi pada variabilitas respons kekeringan di berbagai lokasi, bahkan untuk genotipe yang sama (Zas et al. 2020 ). Misalnya, lokasi dengan zona perakaran yang dalam dapat mendukung genotipe yang memprioritaskan pertumbuhan akar dan mampu mengakses air yang disimpan dari lapisan tanah yang lebih dalam, sementara tegakan yang rapat mungkin mendukung pohon dengan pertumbuhan cepat di atas tanah yang dapat dengan cepat mengalahkan tetangganya (Kozovits et al. 2005 ; Warren et al. 2005 ). Lebih jauh lagi, bagaimana pohon bereaksi terhadap satu kejadian kekeringan dapat memengaruhi bagaimana mereka bereaksi terhadap kejadian kekeringan berikutnya, yang menimbulkan dampak lingkungan tambahan dengan adanya respons yang berkorelasi. Jika kejadian kekeringan terjadi pada frekuensi tinggi, efek kumulatif dapat mendevaluasi penggunaan indeks ketahanan tunggal sebagai sifat respons kekeringan. Meskipun terpapar pada dua kejadian kekeringan serupa di empat lokasi uji keturunan dalam satu wilayah pembiakan (yaitu, kondisi lingkungan yang “seragam”), kami mengamati korelasi rendah antara lokasi atau kejadian kekeringan (Tabel 1 , Tabel S1 ), mirip dengan Zas et al. ( 2020)), di mana respons yang berbeda diamati tergantung pada lokasi atau kejadian kekeringan. Dengan temuan ini, kami mengusulkan indeks penurunan baru yang dapat mengurangi efek kumulatif interaksi antara beberapa kejadian kekeringan berturut-turut dan memberikan estimasi respons kekeringan yang lebih andal dalam uji coba tipe kebun umum yang berusia sama. Indeks penurunan kami menunjukkan estimasi heritabilitas yang lebih tinggi dan lebih stabil di seluruh lokasi daripada indeks respons kekeringan klasik (Tabel 3 ), dan menunjukkan korelasi tertinggi antar lokasi (Tabel S1 ). Penurunan tersebut juga merupakan salah satu dari dua indeks respons kekeringan yang menunjukkan adaptasi signifikan terhadap lingkungan lokal pohon induk (Gambar 6 ).
Beberapa studi telah menunjukkan adaptasi lokal terhadap kekeringan pada konifer menggunakan sifat dendrokronologis (Depardieu et al. 2020 ; Montwé et al. 2016 ) atau pendekatan lain (de la Mata et al. 2014 ; Matías et al. 2014 ; Voltas et al. 2008 ). Studi-studi ini, yang menunjukkan adaptasi lokal, telah digunakan untuk mengadvokasi migrasi berbantuan antara wilayah perkembangbiakan sebagai alat untuk mencapai hutan yang beradaptasi dengan kekeringan (Aitken dan Whitlock 2013 ; Pedlar et al. 2012 ). Dalam studi kami, kami mengamati adaptasi lokal famili pinus lodgepole yang berasal dari dataran rendah yang menunjukkan ketahanan yang lebih baik dan penurunan yang lebih rendah daripada famili dari dataran tinggi. Mendeteksi adaptasi lokal dalam jarak 60 km dan hanya 300 m di atas permukaan laut merupakan hal yang luar biasa, mengingat variasi lingkungan yang ditangkapnya relatif rendah, dibandingkan dengan penelitian lain tentang adaptasi lokal (Depardieu et al. 2020 ; Montwé et al. 2016 ; Zas et al. 2020 ). Hasil-hasil ini menggarisbawahi potensi pemindahan benih dari daerah yang lebih kering ke daerah yang lebih basah untuk meningkatkan respons kekeringan dari suatu sumber benih, bahkan dalam satu wilayah perkembangbiakan, ketika migrasi bantuan dari luar wilayah perkembangbiakan tidak diinginkan.
5 Kesimpulan
Hasil kami menunjukkan bahwa respons kekeringan jangka pendek merupakan pendorong utama kinerja jangka panjang pohon yang terpapar berbagai peristiwa kekeringan. Dengan peningkatan frekuensi kekeringan yang diantisipasi, pohon dengan ketahanan kekeringan cenderung tumbuh lebih cepat, berbeda dengan trade-off yang umum diamati antara ukuran dan respons kekeringan. Lebih jauh, model persamaan struktural mengungkapkan bahwa efek negatif infeksi jamur, dikombinasikan dengan persaingan, dapat memengaruhi respons kekeringan pohon.
Ciri-ciri dendrokronologis menjanjikan untuk pemilihan genotipe yang beradaptasi terhadap kekeringan. Kami menemukan bukti adanya kontrol genetik dalam respons pertumbuhan terhadap kekeringan, dengan estimasi heritabilitas sedang dalam beberapa indeks respons kekeringan, yang sebagian disebabkan oleh adaptasi lokal terhadap lingkungan yang lebih kering. Namun, terdapat variabilitas substansial dalam respons pohon dari famili yang sama terhadap kejadian kekeringan yang berbeda di berbagai lokasi pertumbuhan.
Terakhir, kami memperkenalkan indeks penurunan baru, yang memberikan indikator respons kekeringan yang lebih baik dibandingkan dengan indeks dendrokronologi tradisional. Indeks baru ini berguna ketika beberapa peristiwa kekeringan terjadi dalam waktu singkat, karena indeks ini menangkap efek jangka panjang pada tren pertumbuhan. Indeks penurunan memiliki heritabilitas yang lebih tinggi dan menunjukkan korelasi yang lebih kuat dengan sifat pertumbuhan, sehingga menjadikannya sifat kandidat yang menjanjikan untuk pemilihan genotipe yang beradaptasi terhadap kekeringan dalam program perbaikan pohon.