
Abstrak
Rekrutmen tanaman dibentuk oleh fungsi di seluruh tahap benih dan kecambah. Karena morfologi benih dan perkecambahan secara langsung memengaruhi paparan kecambah terhadap sumber daya dan lingkungan, kedua tahap ini dapat dihubungkan melalui sinergi sifat dan pertukaran yang mengoordinasikan fungsi melalui ontogeni awal. Namun, berbagai sifat yang memengaruhi respons lingkungan pada setiap tahap ontogenetik jarang dieksplorasi secara bersamaan untuk memahami dimensionalitas potensial dari ceruk rekrutmen fungsional.
Kami meneliti kovariasi di antara 13 sifat benih dan bibit yang terkait dengan toleransi stres, laju perkecambahan atau pertumbuhan, respons cahaya, respons suhu, dan fungsi lain untuk 49 spesies yang ditemukan di padang rumput semi-kering. Dengan menggunakan ordinasi yang diinformasikan secara filogenetik dan analisis klaster, kami menanyakan bagaimana kovariasi sifat di berbagai tahap dan fungsi ontogenetik membentuk dimensionalitas ceruk rekrutmen fungsional.
Dua dimensi sifat pertama yang diidentifikasi pada tahap benih dan kecambah yang terpisah selaras, menyediakan beberapa dasar untuk koordinasi ontogenetik selama perekrutan. Sifat morfologi yang mencerminkan toleransi stres terkait ukuran (yaitu massa benih dan kecambah) membentuk fondasi terkuat untuk koordinasi lintas tahap, berbagi ikatan dengan sifat yang mencerminkan respons cahaya kecambah (luas daun tertentu), laju pertumbuhan (perpanjangan akar) dan respons suhu benih (misalnya suhu minimum perkecambahan). Kami juga mengamati trade-off yang tidak terduga dalam cara benih dan kecambah dapat menghindari risiko (melalui dormansi) atau menoleransi risiko (melalui investasi akar), masing-masing.
Sebaliknya, respons cahaya benih, laju perkecambahan benih, dan ambang batas suhu minimum benih tidak terkait erat dengan fungsi analog pada tahap lainnya. Kemandirian mereka dapat memperluas dimensionalitas ceruk perekrutan tergantung pada signifikansi fungsional sifat-sifat ini di lapangan.
Sintesis . Tahap benih dan kecambah dicirikan oleh beberapa dimensi fungsi yang independen, tetapi koordinasi ontogenetik dapat memoderasi dimensionalitas relung rekrutmen fungsional yang semakin meningkat karena berbagai sifat yang lebih luas dieksplorasi bersama. Pada saat yang sama, sifat fisiologis yang terkait dengan respons lingkungan tampak kurang terhubung dengan sifat lain dan dapat memperumit dinamika rekrutmen spasiotemporal. Baik aspek fungsi yang independen maupun terkoordinasi yang diamati di sini layak dieksplorasi di berbagai spesies, sifat, dan lingkungan yang lebih luas untuk memahami dimensionalitas penuh relung rekrutmen fungsional.
1. PENDAHULUAN
Rekrutmen tanaman merupakan kemacetan demografis dengan implikasi besar bagi konservasi dan pemulihan spesies (Albrecht et al., 2019 ; Shackelford et al., 2021 ). Teori ekologi memprediksi bahwa spesies yang hidup berdampingan harus menunjukkan strategi benih dan semai yang beragam (Grubb, 1977 ), yang mengarah pada variasi spasiotemporal dalam rekrutmen yang dapat membentuk dinamika komunitas (Eckhoff et al., 2023 ; Moreno-de las Heras et al., 2016 ). Pada saat yang sama, bidang ekologi fungsional menunjukkan bahwa kita dapat mengkarakterisasi variasi ini dengan mengidentifikasi sifat benih dan semai terkait yang mencerminkan perbedaan dalam fungsi (yaitu bagaimana spesies beroperasi atau merespons konteks lingkungan); bersama-sama, dimensi sifat terkoordinasi ini menyediakan sarana untuk menangkap strategi multidimensi untuk rekrutmen (ceruk rekrutmen fungsional, sensu Larson et al., 2023 ; lihat juga Laughlin, 2014 ). Namun, meskipun ada peningkatan upaya untuk mendeskripsikan variasi sifat pada tahap ontogenetik yang lebih awal (misalnya pada semai, Barton, 2024 ; Winkler et al., 2024 ; atau benih, Saatkamp et al., 2019 ), lebih sedikit penelitian yang mengintegrasikan fungsi lintas tahap benih dan semai sebagai pemengaruh bersama rekrutmen (tetapi lihat, misalnya Gardarin et al., 2016 ; Lamont & Groom, 2013 ). Di sini, kami mengeksplorasi sifat yang mewakili aspek analog dari fungsi pada tahap benih dan semai untuk memahami bagaimana penyelarasan atau independensi mereka membentuk ceruk rekrutmen fungsional (Gambar 1 ).
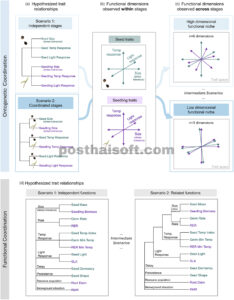
GAMBAR 1
Kami menggunakan variasi sifat benih dan bibit untuk menilai (a–c) koordinasi ontogenetik dan (d) koordinasi fungsional selama perekrutan. Benih dan bibit mewakili tahap morfofisiologis unik selama perekrutan yang mungkin dipengaruhi oleh variasi dalam sifat yang berbeda yang sesuai dengan fungsi analog (misalnya toleransi stres, suhu, dan respons cahaya). (a) Fungsi analog mungkin independen di seluruh tahap ontogenetik (panel atas) atau terkoordinasi sedemikian rupa sehingga fungsi selama satu tahap secara dekat memprediksi fungsi selama tahap lain (panel bawah). (b) Beberapa dimensi fungsional independen dapat terjadi dalam tahap individu, terlepas dari koordinasi ontogenetik (misalnya seperti yang diungkapkan oleh penahbisan sifat rata-rata spesies). Namun, (c) tingkat koordinasi ontogenetik memengaruhi jumlah dimensi fungsional yang diamati di seluruh tahap, dengan fungsi yang tidak terkait di seluruh ontogeni menghasilkan relung fungsional berdimensi tinggi (panel atas) dan sinergi fungsional di seluruh ontogeni menghasilkan relung fungsional berdimensi lebih rendah (panel bawah). (d) Dimensionalitas relung perekrutan bergantung tidak hanya pada koordinasi ontogenetik dalam fungsi tertentu (misalnya toleransi stres atau laju perkembangan), tetapi juga struktur keterkaitan di seluruh fungsi. Dalam studi ini, setiap sifat benih (teks hijau) dan sifat bibit (teks ungu) diasumsikan mewakili fungsi tertentu berdasarkan literatur (teks hitam; Saatkamp et al., 2019 , Winkler et al., 2024 ). Fungsi yang berbeda mungkin sepenuhnya independen (panel kiri) atau menunjukkan tingkat keterkaitan yang jelas dan berbeda (panel kanan), atau beberapa skenario di antaranya. Sementara ordinasi mengungkapkan dimensi fungsional yang sepenuhnya independen, analisis klaster (di sini digambarkan sebagai dendrogram) dapat mengungkapkan struktur keterkaitan yang lebih bernuansa di antara sifat dan fungsi (misalnya, di panel kanan, metrik ukuran dan laju lebih erat kaitannya daripada sifat respons suhu). Lihat Tabel 1 untuk deskripsi sifat lengkap.
Perbedaan morfofisiologis yang jelas memisahkan benih dan bibit (Gambar 2 ), dengan sifat unik yang mendorong respons dan fungsi lingkungan pada setiap tahap (Tabel 1 ). Pada tahap benih, beberapa sifat dapat memengaruhi kapan dan di mana perkecambahan terjadi sebagai respons terhadap lingkungan. Selain sifat yang mencerminkan perbedaan besar dalam tingkat perkecambahan dan penundaan (misalnya dormansi benih), waktu perkecambahan juga dipengaruhi oleh sifat yang mencerminkan respons terhadap faktor lingkungan tertentu seperti cahaya dan suhu (misalnya Carta et al., 2022 ; Zhang et al., 2022 ). Ukuran dan bentuk benih juga dapat memengaruhi paparan dan respons terhadap bahaya lingkungan melalui hubungan dengan penguburan benih dan persistensi di tanah (melalui benih kecil dan bulat) serta toleransi stres selama pembentukan (melalui benih yang lebih besar; Lamont & Groom, 2013 ; Long et al., 2015 ; Tóth et al., 2022 ). Setelah perkecambahan terjadi, bibit segera rentan terhadap bahaya lingkungan, namun sifat dan strategi mungkin berbeda dari yang dipahami pada tanaman dewasa (Barton, 2024 ). Pada saat dewasa, strategi independen dari konstruksi daun, strategi akar, dan ukuran diharapkan memengaruhi perolehan sumber daya cahaya dan bawah tanah (misalnya Weigelt et al., 2021 ). Namun, pada tahap pembibitan, ukuran membedakan akses ke sumber daya di skala spasial yang lebih kecil, dan variasi dalam laju pertumbuhan dan respons suhu (yang membatasi pertumbuhan) juga dapat memiliki dampak awal yang penting pada akses sumber daya (misalnya Mummey et al., 2016 ).
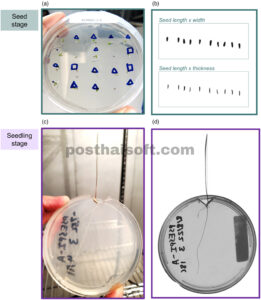
GAMBAR 2
Kami mengukur sifat-sifat pada tahap benih (a, b) dan tahap bibit (c, d), menggunakan uji perkecambahan dan pertumbuhan (a, c) serta perangkat lunak pemindaian untuk menangkap morfologi (b, d).
TABEL 1. Deskripsi sifat benih dan bibit yang diukur dalam penelitian ini.
Sifat-sifat Fungsi yang dihipotesiskan Keterangan Catatan
Benih
Massa benih Toleransi stres terkait ukuran Rata-rata massa kering per biji (mg) transformasi ln
Bentuk biji Ketahanan di dalam tanah Varians ukuran panjang, lebar dan ketebalan benih berskala
Tingkat kuman Tingkat perkembangan (perkecambahan) [Tingkat perkecambahan] Kebalikan dari waktu perkecambahan (hari -1 ) untuk persentil ke-10, dalam kondisi cahaya terbaik dan pemecahan dormansi transformasi ln
Indeks suhu benih Respon sementara [Indeks suhu benih] Perbedaan absolut dalam proporsi benih yang berkecambah antara 18°C dan 6°C, dalam kondisi cahaya terbaik dan pemecahan dormansi
Kuman. suhu min. Respon sementara [Suhu minimum perkecambahan] Estimasi suhu minimum (°C) untuk perkecambahan persentil ke-10 benih berdasarkan x-intercept regresi linier antara suhu dan laju perkecambahan, dalam kondisi cahaya terbaik dan pemecahan dormansi.
Indeks cahaya benih Respon cahaya Perbedaan absolut dalam proporsi benih yang berkecambah antara kondisi terang dan gelap, pada suhu 18°C dalam kondisi terbaik untuk memutus dormansi
Indeks dormansi benih Penghindaran stres atau risiko Proporsi benih yang layak yang tidak berkecambah pada suhu 18°C dalam kondisi cahaya terbaik
Waktu termal kuman Respon sementara. Akumulasi termal yang diperlukan untuk perkecambahan benih persentil ke-10, berdasarkan kemiringan terbalik regresi linier antara suhu dan tingkat perkecambahan (°C hari) Transformasi ln; Redundan dengan laju germ., r = −0.89
Bibit
Biomassa bibit (hari ke-1 ) Toleransi stres terkait ukuran Biomassa kering bibit akhir pada suhu 18°C distandarisasi berdasarkan jumlah hari tumbuh (mg hari -1 ) transformasi ln
Bahasa Indonesia: RER Tingkat perkembangan (pertumbuhan) [Laju pemanjangan akar] Perbedaan antara panjang akar akhir dan awal pada suhu 18°C distandarisasi dengan jumlah hari tumbuh (cm hari −1 ) transformasi ln
RER Suhu Min. Respon sementara [Suhu minimum RER] Estimasi suhu minimum (°C) untuk pemanjangan akar berdasarkan intersep x regresi linier antara suhu dan RER
Persyaratan Layanan Akuisisi cahaya [Luas daun spesifik] Luas daun per massa daun kering pada suhu 18°C (cm 2 g −1 ). Jaringan ‘daun’ membentuk kotiledon pada dikotil, karena daun sejati belum muncul. transformasi ln
Akar diam. Akuisisi sumber daya bawah tanah [Diameter akar] Diameter akar rata-rata pada suhu 18°C (mm); Berhubungan positif dengan simbiosis mikroba untuk perolehan sumber daya (Ma et al., 2018 ) transformasi ln
Rasio massa akar Akuisisi (alokasi) sumber daya bawah tanah Rasio biomassa akar kering terhadap total biomassa bibit pada suhu 18°C (mg mg −1 )
Waktu termal RER Respon sementara Akumulasi termal yang diperlukan untuk pemanjangan akar, berdasarkan kemiringan terbalik regresi linier antara suhu dan RER (°C hari cm −1 ) Transformasi ln; Redundan dengan RER, r = 0,92
Bahasa Inggris Akuisisi sumber daya bawah tanah [Panjang akar spesifik] Panjang akar per massa kering pada suhu 18°C (m g −1 ) Transformasi ln; Redundan dengan diameter akar, r = −0.93
Catatan : ‘Ciri-ciri’ merujuk pada atribut spesies yang diukur secara tepat yang digunakan sebagai indikator langsung atau proksi tidak langsung untuk ‘fungsi’, yang menangkap perbedaan dalam cara spesies beroperasi atau merespons konteks lingkungan.
Premis utama ekologi fungsional adalah bahwa pertukaran sifat dan sinergi—yang diperlukan oleh hubungan fungsional atau kendala fisik—mengarah ke subset dimensi fungsional yang lebih kecil yang merangkum perbedaan utama di antara spesies. Misalnya, penelitian biasanya mengidentifikasi empat hingga enam dimensi yang menangkap berbagai aspek fungsi pada tanaman dewasa (misalnya Laughlin, 2014 ; Weigelt et al., 2021 ). Jumlah dimensi fungsional yang muncul selama perekrutan akan bergantung pada tingkat koordinasi antara tahap ontogenetik (yaitu benih dan bibit) serta fungsi yang berbeda pada tahap-tahap ini (Gambar 1 ). Perbedaan dalam toleransi stres, laju perkecambahan/pertumbuhan dan respons lingkungan pada setiap tahap masing-masing dapat terdiri dari dimensi fungsional yang independen (Gambar 1a,c , panel atas). Namun, koordinasi ontogenetik diharapkan ketika respons lingkungan pada tahap benih pada dasarnya membentuk paparan bibit terhadap kondisi lingkungan (misalnya cahaya atau suhu, Donohue et al., 2010 ), atau ketika hubungan fisik mendasar terjadi (misalnya benih besar menghasilkan bibit yang lebih besar, Lamont & Groom, 2013 ). Jika sifat yang mewakili fungsi analog berkorelasi di seluruh tahap ontogenetik, ini dapat menyederhanakan ceruk rekrutmen fungsional menjadi lebih sedikit dimensi (Gambar 1a,c , panel bawah). Namun, karena sifat dieksplorasi dalam kaitannya dengan fungsi yang lebih luas, beberapa di antaranya mungkin unik untuk tahap ontogenetik tertentu, dimensionalitas ceruk rekrutmen juga akan bergantung pada struktur keterkaitan di antara fungsi (Gambar 1d dan keterangan).
Bahasa Indonesia: Dalam studi ini, kami mengeksplorasi variasi sifat benih dan bibit (Tabel 1 ) di 49 spesies yang ditargetkan untuk digunakan dalam restorasi berbasis benih di padang rumput semi-kering AS. Spesies ini umumnya ditargetkan karena diinginkan untuk fungsi ekosistem seperti konservasi habitat satwa liar atau pengendalian erosi, atau karena mereka umum di rentang geografis yang luas. Karena potensi penggunaannya dalam restorasi, sifat benih dan bibit dari spesies ini juga menarik untuk aplikasi, misalnya, untuk membantu memilih spesies yang lebih mungkin untuk merekrut dalam kondisi yang berbeda (Larson et al., 2023 ). Ada bukti yang berkembang bahwa ontogeni awal—sekitar waktu perkecambahan dan munculnya bibit—merupakan pendorong penting variasi interspesifik dalam perekrutan tahun pertama, dan bahwa sifat-sifat awal di seluruh tahap ini dapat menginformasikan pola-pola ini (misalnya Larson et al., 2021 ; Zirbel & Brudvig, 2020 ). Pada saat yang sama, informasi sifat yang terkait dengan perkecambahan dan perekrutan masih terbatas untuk spesies restorasi yang ditargetkan (Ladouceur et al., 2018 ), seperti halnya panduan tentang sifat mana yang akan diukur.
Untuk mengeksplorasi ceruk rekrutmen fungsional, pertama-tama kami mengevaluasi sifat benih dan kecambah secara terpisah untuk mengkarakterisasi keterkaitan sifat dan dimensionalitas dalam setiap tahap. Kami kemudian mengevaluasi tahap-tahap tersebut bersama-sama, menanyakan bagaimana koordinasi sifat di berbagai tahap dan fungsi ontogenetik membentuk dimensionalitas ceruk rekrutmen. Kami berharap menemukan koordinasi antara tahap-tahap ontogenetik tempat analog fungsional ada (yaitu sifat benih dan kecambah dengan hubungan yang diharapkan serupa dengan toleransi stres, laju, suhu atau respons cahaya). Namun, kami juga berharap mengidentifikasi dimensi independen yang mencerminkan aspek unik dari fungsi pada setiap tahap—misalnya, jika strategi penundaan perkecambahan pada tahap benih (misalnya dormansi dan persistensi dalam tanah) tidak memiliki fungsi yang setara pada tahap kecambah. Hal ini dapat menyebabkan ceruk fungsional berdimensi tinggi di seluruh tahap, menandakan potensi strategi rekrutmen yang beragam di antara spesies yang hidup berdampingan. Sementara 49 spesies ini merupakan bagian kecil dari total keanekaragaman tumbuhan di padang rumput semi-kering ini, mereka mencakup beberapa famili tumbuhan terbesar (Asteraceae, Fabaceae, Poaceae). Banyak studi sifat memperhitungkan keterkaitan filogenetik dengan tujuan mengidentifikasi hubungan sifat dengan peran adaptif dalam lingkungan kontemporer (dibandingkan dengan kesamaan sisa antara spesies yang berkerabat dekat). Namun, tidak semua studi sifat menggunakan pendekatan filogenetik, dan kovariasi sifat yang terkait dengan sejarah evolusi masih dapat relevan secara fungsional jika evolusi filogenetik dan relung telah terjadi bersamaan (Westoby et al., 2023 ). Akibatnya, kami menganalisis pola dengan dan tanpa informasi filogenetik untuk memperluas inferensi dan meningkatkan keterbandingan dengan studi lain.
2 BAHAN DAN METODE
2.1 Studi spesies
Kami berkonsultasi dengan para manajer lokal, peneliti dan spesialis bahan tanaman untuk memilih 47 spesies studi asli yang menjadi target restorasi skala besar di padang rumput semi-kering dan lahan semak belukar AS bagian barat yang tersedia dari sumber komersial atau diidentifikasi untuk upaya produksi benih di masa mendatang (Lampiran S1 : Tabel A1, lihat juga Gambar 3 ). Meskipun spesies bervariasi dalam distribusi geografis, semua menghuni sistem yang dicirikan oleh musim dingin yang dingin dan basah serta musim panas yang hangat dan kering. Spesies asli meliputi forb tahunan dan abadi, rumput musim dingin abadi (C3) dan musim panas (C4), dan semak dari 12 famili tanaman. Famili yang paling umum adalah Asteraceae, Fabaceae dan Poaceae yang mencerminkan kontribusi relatif mereka terhadap kekayaan spesies di Amerika Serikat bagian barat, dan secara global (Christenhusz & Byng, 2016 ). Kami juga menyertakan dua spesies rumput eksotis yang sering bersaing dengan tanaman asli selama restorasi berbasis benih, rumput gandum jambul abadi ( Agropyron cristatum ) dan cheatgrass tahunan ( Bromus tectorum ) sebagai perbandingan (total spesies, n = 49).
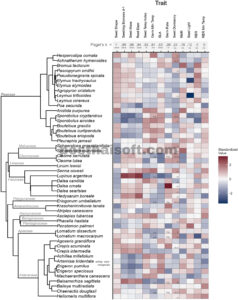
GAMBAR 3
Peta panas nilai sifat rata-rata spesies. Sifat (yaitu kolom) disusun menurut sinyal filogenetik dan spesies (yaitu baris) disusun menurut keterkaitan filogenetik. Famili tumbuhan ditampilkan sebagai simpul dalam pohon filogenetik. Dalam peta panas, kami menunjukkan nilai sifat yang diperhitungkan atau disubstitusi untuk laju perkecambahan dan suhu minimum, dan perlakuan pemutusan dormansi yang diterapkan terbaik untuk dormansi benih. Imp ., nilai sifat yang diperhitungkan; sub ., nilai sifat yang disubstitusi; st ., stratifikasi; sc , skarifikasi; st-sc , stratifikasi dan skarifikasi. Nilai signifikansi untuk sinyal filogenetik: * p < 0,05, ** p < 0,01, *** p < 0,001, ns, tidak signifikan.2.2 Pengukuran sifat Kami mengukur 13 sifat benih dan bibit (Tabel 1 ). Untuk melakukan ini, kami mengembangkan protokol penyaringan standar untuk memfasilitasi penyederhanaan dan kelayakan. Standarisasi ini terkadang membatasi kesimpulan, khususnya berkenaan dengan kondisi perkecambahan dan pemutusan dormansi, yang dapat bervariasi dalam berbagai cara yang rumit di antara spesies. Namun, pendekatan ini memungkinkan kami untuk menangkap perbedaan utama di antara lusinan spesies dengan waktu dan sumber daya yang terbatas. Kami memberikan ikhtisar tentang metode yang terkait dengan sifat di sini, dan protokol eksperimen terperinci di Lampiran S2 . 2.3 Ciri-ciri morfologi benih Untuk memperkirakan massa biji , kami membuang apendiks eksternal (misalnya duri, pappus, sekam, lemma, dan palea) lalu menghitung, mengeringkan (80°C selama 3 hari), dan menimbang tiga ulangan benih per spesies (15–100 biji, tergantung ukuran). Untuk memperkirakan bentuk biji , kami memindai gambar 10 ulangan dispersi per spesies (struktur eksternal biji dipertahankan kecuali duri panjang, Gambar 2b ), lalu memperkirakan panjang, lebar, dan tebal dengan perangkat lunak otomatis (traitor, Dayrell et al., 2023 ). Kami mengukur bentuk biji sebagai metrik varians, dengan nilai yang lebih kecil menunjukkan bentuk yang lebih bulat dan nilai yang lebih besar menunjukkan bentuk yang memanjang atau pipih (Tabel 1 ). 2.4 Sifat perkecambahan dan dormansi biji Kami melakukan serangkaian uji perkecambahan benih untuk menilai persyaratan dormansi (skarifikasi, stratifikasi, keduanya atau tidak keduanya), persyaratan cahaya (terang atau gelap) dan sensitivitas suhu (respons pada 6°C, 12°C dan 18°C). Kami memilih suhu ini berdasarkan pengalaman sebelumnya dengan beberapa spesies yang menunjukkan bahwa mereka berada dalam kisaran antara suhu perkecambahan minimum dan optimal (Larson et al., 2016 ), meskipun persyaratan yang lebih hangat untuk beberapa spesies mendukung rentang suhu yang lebih luas dalam percobaan di masa depan. Terlepas dari perlakuan, setiap uji perkecambahan terdiri dari desain yang sama: Kami membuat enam replikasi per spesies yang terdiri dari 20 benih yang diisi masing-masing (atau 10, untuk spesies yang bijinya terbatas), didistribusikan secara merata pada agar bebas nutrisi 1% dalam cawan Petri 90 mm yang disterilkan yang disegel dengan parafilm dan ditempatkan di ruang lingkungan yang terkontrol (SG2-22, Hoffman Manufacturing, Corvallis, OR, AS; Gambar 2a ). Kami menghitung perkecambahan harian selama 7 hari, kemudian setiap 2–3 hari selama minimal 28 hari total (sebagian besar uji coba pada suhu 18°C) dan hingga 49 hari (sebagian besar uji coba pada suhu 6°C dan 12°C). Kami mensterilkan benih dengan merendamnya dalam larutan pemutih 0,5% dengan surfaktan 1%, dan pada akhir setiap uji coba, menggunakan uji potong untuk menilai viabilitas benih yang tersisa. Kami mengikuti desain studi hierarkis untuk menerapkan perlakuan perkecambahan yang tepat (dormansi, cahaya dan/atau suhu) untuk setiap spesies (lihat diagram alir di Lampiran S2 : Gambar B1). Kami pertama-tama mengevaluasi perkecambahan dalam kondisi terang (fotoperiode 12 jam) dan gelap pada suhu perkecambahan terhangat (18°C) tanpa perlakuan pemecah dormansi. Spesies dengan persentase perkecambahan rata-rata >70% (disesuaikan dengan viabilitas) diklasifikasikan sebagai tidak dorman ( n = 22). Kami menguji perlakuan untuk memecah dormansi fisik (skarifikasi) dan fisiologis (stratifikasi) untuk spesies yang tersisa ( n = 27). Untuk spesies dalam famili yang dikenal dengan dormansi fisik yang luas (Fabaceae, Malvaceae; n = 10), kami hanya menguji skarifikasi. Untuk spesies lain, kami menguji skarifikasi, stratifikasi dan kombinasinya. Kami melakukan skarifikasi benih dengan cara menggores setiap benih dengan silet halus. Kami membuat stratifikasi benih dengan menaruhnya di antara kertas isap yang dibasahi (~47% kadar air gravimetrik) dalam cawan Petri pada suhu 3°C selama 6 minggu dalam keadaan gelap. Kami membuat skarifikasi benih sebelum stratifikasi untuk perlakuan kombinasi. Setelah perlakuan, kami menumbuhkan benih dengan metode yang sama seperti percobaan dormansi awal (lihat di atas).
Kami menggunakan hasil ini untuk mengidentifikasi kondisi cahaya dan pemutusan dormansi terbaik untuk setiap spesies. Kami melakukan uji ANOVA terpisah untuk setiap spesies dengan perlakuan cahaya atau interaksi perlakuan cahaya dan pemutusan dormansi sebagai efek tetap, dan persentase perkecambahan total sebagai respons. Karena lot benih bervariasi dalam proporsi benih yang awalnya layak, kami mengukur perkecambahan total berdasarkan kelangsungan hidup lot benih, yang diperkirakan sebagai persentase maksimum benih yang dapat berkecambah atau layak (berdasarkan uji potong) untuk setiap spesies, secara rata-rata, di bawah perlakuan perkecambahan apa pun. Kami menetapkan ‘gelap’ sebagai kondisi cahaya terbaik untuk setiap spesies kecuali perlakuan ‘terang’ secara signifikan meningkatkan perkecambahan total. Demikian pula, jika beberapa metode pemutusan dormansi secara statistik setara sebagai perlakuan ‘terbaik’, kami menetapkan perlakuan yang paling sederhana (urutan: tidak ada > skarifikasi > stratifikasi > skarifikasi dan stratifikasi, hasil dalam Lampiran S1 : Tabel A1). Kami kemudian menerapkan perlakuan cahaya dan pemutusan dormansi terbaik untuk uji perkecambahan pada suhu 6°C, 12°C, dan jika perlu, 18°C (diulang hanya untuk spesies yang terstratifikasi). Kami mengikuti protokol yang sama seperti di atas tetapi menyesuaikan perlakuan stratifikasi dengan mengurangi periode 6 minggu menjadi 2 minggu untuk spesies yang sebelumnya telah berkecambah selama stratifikasi ( n = 5 dari 10 spesies) dan menggandakannya menjadi 12 minggu untuk spesies dengan perkecambahan <10% di bawah stratifikasi 6 minggu ( n = 2). Kami juga mengubah media stratifikasi menjadi 1% agar (dari kertas isap) untuk mempertahankan tingkat kelembapan yang konsisten untuk pemodelan termal. Kami menggunakan data dari uji perkecambahan ini untuk memperkirakan beberapa indeks yang terkait dengan dormansi, cahaya, dan respons suhu. Semua indeks didasarkan pada persentase perkecambahan yang dirata-ratakan di seluruh replikasi perlakuan dan diskalakan berdasarkan viabilitas semai (yaitu viabilitas semai rata-rata maksimum yang diamati di bawah setiap perlakuan perkecambahan). Kami memperkirakan indeks cahaya benih sebagai perbedaan persentase perkecambahan total rata-rata antara perlakuan terang dan gelap di bawah kondisi pemutusan dormansi ‘terbaik’ pada suhu 18°C (nilai positif dan negatif menunjukkan lebih banyak perkecambahan dalam terang atau gelap, masing-masing). Kami memperkirakan indeks dormansi benih sebagai proporsi rata-rata benih yang layak yang tidak berkecambah di bawah kondisi cahaya ‘terbaik’ pada suhu 18°C ketika tidak ada perlakuan pemutusan dormansi yang diterapkan (nilai yang lebih tinggi menunjukkan lebih banyak dormansi). Akhirnya, kami memperkirakan indeks suhu benih sebagai perbedaan persentase perkecambahan total rata-rata antara 18°C dan 6°C, dengan kondisi cahaya ‘terbaik’ dan pemutusan dormansi diterapkan (nilai positif menunjukkan lebih banyak perkecambahan pada suhu hangat). Kami juga memperkirakan beberapa sifat berdasarkan waktu perkecambahan. Kami menggunakan uji perkecambahan pada 6°C, 12°C dan 18°C untuk memperkirakan suhu minimum perkecambahan , yang didefinisikan sebagai suhu terendah di mana perkecambahan diharapkan terjadi untuk persentil perkecambahan yang dipilih. Kami menggunakan persentil ke-10 (mencerminkan respons benih yang lebih awal dan lebih kuat), karena banyak spesies tidak mencapai persentase yang lebih tinggi pada suhu yang lebih dingin. Untuk setiap spesies dan suhu, kami membuat kurva perkecambahan kumulatif yang dirata-ratakan di semua enam replikasi (diskalakan oleh perkiraan viabilitas benih setiap spesies). Dengan menggunakan interpolasi linier antara langkah waktu yang diamati, kami memperkirakan waktu perkecambahan untuk setiap persentil ke-5, kemudian mengambil kebalikan dari nilai-nilai ini untuk mendapatkan tingkat perkecambahan (hari -1 ). Mengekstrak tingkat perkecambahan untuk persentil ke-10, kami menyesuaikan regresi linier untuk setiap spesies antara suhu dan tingkat perkecambahan, kemudian memperkirakan suhu minimum perkecambahan sebagai intersep-x. Kami juga memperkirakan waktu termal perkecambahan sebagai kemiringan terbalik dari hubungan yang dimodelkan; ini menggambarkan akumulasi suhu di atas ambang batas suhu minimum setiap spesies (dalam derajat hari, °C hari) yang diperlukan untuk persentil ke-10 benih untuk berkecambah. Pendekatan pemodelan mengikuti Larson et al. ( 2016 ) dan umumnya menghasilkan estimasi kecocokan model linier R 2 > 0,90 (lihat Lampiran S1 : Gambar A1 untuk detailnya). Akhirnya, kami mengukur laju perkecambahan sebagai kebalikan dari waktu hingga 10% perkecambahan pada 18°C dengan cahaya ‘terbaik’ dan kondisi pemecahan dormansi (yaitu laju perkecambahan dalam kondisi yang menguntungkan). Kami membuat beberapa pengecualian terhadap pendekatan ini untuk spesies yang terstratifikasi, yang dijelaskan di samping model regresi suhu dalam Lampiran S1 : Gambar A1.
2.5 Ciri-ciri perkembangan bibit
Kami mengkarakterisasi morfologi bibit awal dan sensitivitas terhadap suhu dengan lima bibit ulangan pada masing-masing dari tiga suhu (6°C, 12°C dan 18°C; tumbuh terpisah dari uji perkecambahan yang dijelaskan di atas). Setiap bibit dipindahkan ke jendela yang dipotong ke sisi sempit cawan Petri steril berukuran 90 mm yang diisi dengan 30 mL agar bebas nutrisi 1% dan disegel dengan parafilm (Gambar 2c ; Quigley et al., 2023 ). Kami mengurangi paparan cahaya akar dengan membungkus setiap cawan dengan foil, kemudian menempatkan cawan secara vertikal di ruang lingkungan yang terkontrol (periode cahaya 12 jam) selama ~1 minggu pertumbuhan (atau 2 minggu untuk beberapa spesies yang tumbuh lambat).
Kami mengambil pemindaian berulang pada setiap bibit pada hari transplantasi (Hari 0) dan hari panen (biasanya Hari 7) menggunakan pemindai flatbed (Gambar 2d , Epson Expression 10000xl). Dari gambar-gambar ini, kami memperoleh panjang dan diameter akar pada setiap titik waktu (WinRhizo v.2.0, Regent Instruments, Ste. Foy, Quebec, Kanada). Setelah pemindaian, kami dengan hati-hati mengeluarkan dan memisahkan setiap bibit menjadi akar, pucuk dan jaringan daun (atau kotiledon). Kami memindai daun segar untuk mengekstrak area (perangkat lunak ImageJ), kemudian mengeringkannya (60°C selama >2 hari) dan menimbang komponen bibit yang terpisah. Kami memperkirakan sifat morfologi bibit mengikuti pedoman yang ditetapkan (Tabel 1 ). Kami terutama menggunakan sifat-sifat yang diukur pada suhu terhangat (18°C), tetapi kami menggunakan nilai laju pemanjangan akar (RER) pada suhu 6°C, 12°C, dan 18°C untuk memperkirakan metrik suhu minimum RER dan waktu termal RER dengan mengikuti pendekatan pemodelan yang sama seperti yang dijelaskan untuk perkecambahan (lihat juga Misra, 1999 ; model regresi ditunjukkan pada Lampiran S1 : Gambar A2).
2.6 Analisis
Sebelum analisis, kami memeriksa distribusi sifat dan mentransformasikan beberapa sifat ke dalam logaritma natural untuk memenuhi persyaratan normalitas multivariat untuk ordinasi (Lampiran S1 : Gambar A3 dan A4). Kami menghilangkan tiga sifat yang redundan secara fungsional dan berkorelasi kuat (| r | > 0,80) dengan sifat lain: waktu termal perkecambahan, waktu termal RER, dan SRL (Tabel 1 ). Meskipun biomassa bibit juga sangat berkorelasi dengan diameter akar dan SLA, kami mempertahankan sifat-sifat ini karena mereka mewakili fungsi yang unik. Kami mengimputasikan nilai sifat yang hilang (suhu minimum perkecambahan, n = 2/49 spesies, laju perkecambahan, n = 6/49, Gambar 3 ) menggunakan pendekatan ordinasi berdasarkan semua sifat benih dan bibit lainnya dalam set data akhir (paket missMDA dalam R; lihat Lampiran S1 : Tabel A2 untuk kutipan perangkat lunak statistik).
2.6.1 Pohon filogenetik dan sinyal
Kami mencocokkan setiap spesies dengan basis data World Plant (Hassler, 2023 ), lalu membuat pohon filogenetik berdasarkan mega-pohon yang dikembangkan oleh Zanne et al. ( 2014 ) dan Smith dan Brown ( 2018 ) menggunakan paket V.Phylomaker2 di R (Jin & Qian, 2022 ). Kami memperkirakan sinyal filogenetik setiap sifat sebagai lambda Pagel, yang dioptimalkan melalui kemungkinan maksimum (paket phytool di R).
2.6.2 Hubungan sifat dalam tahapan
Untuk mengkarakterisasi dimensi fungsional dalam setiap tahap, pertama-tama kami melakukan analisis komponen utama (PCA) secara terpisah untuk sifat benih dan kecambah, dengan dan tanpa mengintegrasikan informasi filogenetik. Tanpa informasi filogenetik, kami melakukan PCA standar (paket vegan di R) dan menggunakan korelasi Pearson bivariat untuk memandu interpretasi. Kami mempertahankan jumlah sumbu PCA berdasarkan aturan Kaiser (paket nFactors di R). Untuk mengintegrasikan filogeni, kami menggunakan PCA filogenetik (paket phytools di R) dengan estimasi kemungkinan maksimum untuk menentukan nilai lambda, berkisar dari 0 (tidak ada sinyal filogenetik) hingga 1 (gerak Brown penuh). Kami juga memperkirakan korelasi sifat bivariat yang diinformasikan secara filogenetik dengan menentukan model kuadrat terkecil filogenetik (PGLS; lambda dioptimalkan dengan kemungkinan maksimum) untuk setiap pasangan sifat dan mengekstraksi koefisien determinasi (Weigelt et al., 2021 ; detail lengkap di Lampiran S1 : Tabel A3).
2.6.3 Hubungan sifat antar tahapan
Bahasa Indonesia: Untuk menilai bagaimana sifat dikoordinasikan di seluruh tahap ontogenetik, kami mengulang PCA di seluruh rangkaian sifat benih dan kecambah seperti yang dijelaskan di atas (dengan dan tanpa filogeni). Kami menggunakan analisis klaster untuk membantu menafsirkan keterkaitan sifat; sementara ordinasi mengidentifikasi rangkaian sifat yang berkorelasi, analisis klaster memberikan nuansa tambahan dengan mengidentifikasi bagaimana subkelompok sifat dalam suatu sumbu terkait dan memungkinkan evaluasi yang lebih jelas tentang harapan bahwa sifat harus mengelompok berdasarkan tahap ontogenetik (yaitu benih versus kecambah) dan/atau fungsi analog (misalnya respons suhu atau cahaya). Mengikuti Maynard et al. ( 2022 ), kami melakukan analisis klaster berdasarkan matriks korelasi sifat yang lengkap dan terinformasi secara filogenetik dan mengidentifikasi jumlah klaster yang optimal menggunakan metode siluet (paket dendextend di R). Kami juga mengeksplorasi hubungan bivariat antara sifat benih dan bibit analog yang mencerminkan toleransi stres terkait ukuran (massa benih dan bibit), laju (laju perkecambahan dan pemanjangan akar), respons suhu (suhu minimum perkecambahan dan RER, indeks suhu benih) dan respons cahaya (indeks cahaya benih dan SLA).
3 HASIL
3.1 Sinyal filogenetik
Kami mendeteksi sinyal filogenetik yang signifikan dalam delapan dari 13 sifat (Gambar 3 ). Secara umum, sifat-sifat yang terkait dengan ukuran dan morfologi sangat terkonservasi secara filogenetik. Di antara sifat-sifat biji, sinyal terkuat terjadi pada bentuk dan massa biji ( λ > 0,98), meskipun metrik suhu juga sangat terkonservasi (suhu minimum kecambah dan indeks suhu biji, λ > 0,80). Kami mengamati sinyal filogenetik yang lemah untuk laju perkecambahan (setelah menerapkan kondisi pemecahan dormansi dan cahaya terbaik; λ = 0,39), tetapi tidak untuk dormansi biji atau indeks cahaya. Di antara sifat-sifat kecambah, biomassa dan diameter akar memiliki sinyal terkuat ( λ > 0,90) diikuti oleh SLA ( λ = 0,63), tetapi kami tidak mendeteksi sinyal signifikan untuk pertumbuhan dan sifat fisiologis (yaitu rasio massa akar [RMR], laju pemanjangan akar [RER] dan suhu minimum RER).
Beberapa sinyal filogenetik tampaknya didorong oleh perbedaan antara famili tumbuhan utama. Misalnya, Poaceae dan Fabaceae masing-masing menunjukkan bentuk biji yang relatif linier dan bulat (Lampiran S1 : Gambar A5B), dan berbeda karena memiliki kecambah yang lebih kecil dan berjaringan tipis atau kecambah yang lebih besar dan berjaringan tebal (Lampiran S1 : Gambar A5D). Sinyal filogenetik lainnya tidak mudah dipisahkan berdasarkan famili tumbuhan, dan hanya sedikit pola terkait famili yang tersisa setelah informasi filogenetik dimasukkan dalam analisis (yaitu PCA filogenetik, Lampiran S1 : Gambar A5A,C,E).
3.2 Fungsi terkoordinasi dalam tahap ontogenetik
3.2.1 Sifat-sifat benih
PCA mengidentifikasi beberapa dimensi independen dalam tahap benih (Gambar 4 ), yang mencerminkan beberapa korelasi kuat di antara sifat-sifat benih (| r | > 0,5, Gambar 5 ; Lampiran S1 : Tabel A3). Kami mempertahankan empat sumbu untuk menggambarkan variasi di tujuh sifat dengan PCA filogenetik (Gambar 4A ) atau tiga sumbu dengan PCA non-filogenetik (Lampiran S1 : Gambar A5; Tabel A4). Dalam kedua analisis, sumbu pertama menunjukkan bahwa spesies dengan massa benih yang lebih besar cenderung memiliki suhu minimum perkecambahan dan nilai indeks suhu benih yang lebih rendah (phylo PC1 = 31,6% variasi, PC1 standar = 32,4%). Namun, pola selanjutnya berbeda berdasarkan pendekatan. Dengan PCA filogenetik, sumbu kedua menunjukkan trade-off antara indeks cahaya benih dan dormansi benih (phylo PC2 = 19,7% variasi), meskipun sifat-sifat ini hanya berkorelasi lemah (PGLS r = −0,21). Sumbu ketiga menangkap variasi dalam tingkat perkecambahan (phylo PC3 = 15% variasi), sedangkan sumbu keempat menangkap variasi tambahan dalam bentuk biji dan indeks cahaya biji (dua sifat yang tidak berkorelasi, PGLS r = -0,05; phylo PC4, 14,7% variasi). Ketika filogeni tidak disertakan, kami mengamati trade-off yang lebih kuat antara bentuk biji dan waktu perkecambahan pada sumbu kedua (PC2 = 23,5% variasi), di mana bentuk biji yang lebih bulat dikaitkan dengan dormansi biji yang lebih besar (Pearson r = -0,37), kebutuhan cahaya biji (Pearson r = -0,27) dan tingkat perkecambahan setelah kondisi pemutusan dormansi/cahaya terbaik diterapkan (Pearson r = -0,34).


Dendrogram yang menunjukkan analisis klaster sifat benih (hijau) dan sifat bibit (ungu) berdasarkan korelasi sifat yang diinformasikan secara filogenetik (lihat Lampiran S1 : Gambar A6 untuk dendrogram berdasarkan korelasi sifat mentah). Korelasi yang diinformasikan secara filogenetik ditunjukkan pada diagonal bawah dari plot gelembung kiri dan korelasi Pearson standar antara nilai sifat mentah ditunjukkan pada diagonal atas. Kami juga menunjukkan pemuatan sifat pada setiap sumbu PCA filogenetik (plot gelembung tengah) dan kekuatan sinyal filogenetik untuk setiap sifat pada plot gelembung kanan. Di semua plot, hitam mewakili nilai positif dan abu-abu mewakili nilai negatif. Desain gambar terinspirasi oleh Maynard et al. ( 2022 ).
3.2.2 Ciri-ciri Bibit
Kami mengamati beberapa korelasi sifat kuat pada tahap pembibitan (Gambar 5 ), dan tiga sumbu menjelaskan sebagian besar variasi di enam sifat pembibitan dengan PCA (Gambar 4B ; Lampiran S1 : Tabel A3). Hasilnya secara umum konsisten antara pendekatan PCA filogenetik dan standar (Lampiran S1 : Gambar A5; Tabel A4), jadi kami menggambarkan pola yang diinformasikan secara filogenetik di sini. Sumbu pertama menangkap hubungan potensial antara toleransi stres terkait ukuran, respons cahaya, dan laju pertumbuhan (PC1 = 44,7% variasi). Secara khusus, spesies dengan biomassa pembibitan yang lebih besar juga memiliki diameter akar yang lebih besar ( r = 0,89), SLA yang lebih rendah ( r = −0,78) dan RER yang lebih cepat ( r = 0,56). Sumbu kedua menangkap variasi tambahan dalam RER dan RMR (PC2 = 24,9% variasi), menunjukkan bahwa spesies yang mengalokasikan lebih banyak biomassa ke akar juga memiliki pemanjangan akar yang lebih cepat ( r = 0,34). Terakhir, suhu minimum RER direpresentasikan pada sumbu ketiga (PC3 = 19,2% variasi) dan memiliki korelasi lemah dengan sifat bibit lainnya.
3.3 Fungsi terkoordinasi di seluruh tahapan ontogenetik
PCA filogenetik dan standar mengidentifikasi pola serupa dalam dua dimensi terkuat dari variasi sifat untuk gabungan sifat biji dan bibit (Lampiran S1 : Gambar A5; Tabel A4), jadi kami hanya menjelaskan hasil yang diinformasikan secara filogenetik di sini (Gambar 4c ). Sumbu pertama menangkap sinergi yang kuat antara massa biji, biomassa bibit dan diameter akar sebagai aspek toleransi stres terkait ukuran (lihat juga Gambar 6a,b ) dan menghubungkan ini dengan metrik laju, cahaya dan suhu. Secara khusus, spesies dengan biji, bibit dan diameter akar yang lebih besar cenderung memiliki RER yang lebih cepat, SLA yang lebih rendah dan nilai suhu minimum perkecambahan dan indeks suhu benih yang lebih rendah (PC1, 34,2% dari variasi). Hubungan ini juga ditangkap oleh analisis klaster (Gambar 5 ), yang mengidentifikasi massa biji, biomassa bibit dan diameter akar sebagai klaster sifat fungsional yang paling rapat dengan hubungan dekat (dalam urutan) dengan sifat SLA, RER dan suhu benih.

Pasangan sifat bivariat yang menunjukkan hubungan antara sifat analog yang diharapkan mewakili fungsi serupa pada berbagai tahap, termasuk (a, b) sifat toleransi stres terkait ukuran, (c, d) sifat respons suhu, (e) metrik laju dan (f) sifat respons cahaya. Di setiap panel, titik-titik diwarnai berdasarkan famili tumbuhan (Asteraceae, Fabaceae, Poaceae atau lainnya), dan garis-garis halus menunjukkan perkiraan hubungan linier untuk setiap kelompok tanpa indikasi signifikansi statistik apa pun. Kami tidak memperkirakan model linier, tetapi melaporkan korelasi yang diinformasikan secara filogenetik (phylo r ) dan korelasi standar (Pearson r ) untuk setiap kombinasi sifat, yang diperkirakan di seluruh spesies. Titik-titik yang digarisbawahi dalam warna hijau tua mewakili dua spesies eksotis yang termasuk dalam analisis
Sumbu PCA kedua menangkap trade-off antara dormansi benih dan RMR ( r = −0,41), sehingga spesies dengan dormansi rendah (yaitu penundaan perkecambahan lebih sedikit) cenderung mengalokasikan lebih banyak ke akar pada tahap pembibitan (PC2 = 15,5% variasi). Dalam analisis klaster, sifat-sifat ini paling dekat dengan yang terwakili pada PC1, karena dormansi rendah dan RMR tinggi juga berasosiasi lemah dengan atribut seperti SLA tinggi dan RER cepat (Lampiran S1 : Tabel A3). Ketika kami tidak memasukkan filogeni, bentuk benih juga dikaitkan dengan sumbu ini (Lampiran S1 : Gambar A5) mengingat korelasinya yang moderat dengan dormansi benih (Pearson r = −0,37). Akhirnya, sumbu ketiga dan keempat dari PCA filogenetik merepresentasikan sifat tunggal dengan ikatan lemah ke fungsi lain, termasuk suhu minimum RER (PC3: 11,5% variasi) dan indeks cahaya benih (PC4: 9,7% variasi).
Meskipun jumlah sumbu yang dipertahankan dalam PCA gabungan ( n = 4) tidak meningkat relatif terhadap tahap benih dan kecambah individu ( sumbu n = 4 dan n = 3, berturut-turut), beberapa sifat tidak terwakili dengan baik oleh sumbu ini, termasuk laju perkecambahan dan bentuk benih. Analisis klaster sifat benih dan kecambah juga menunjukkan bahwa laju perkecambahan benih, indeks cahaya benih, bentuk benih dan suhu minimum RER terkait jauh dengan sifat-sifat lain, meningkatkan jumlah klaster sifat unik ( n = 8, atau sedikit lebih sedikit dengan pendekatan non-filogenetik, Lampiran S1 : Gambar A6). Lebih jauh lagi, ketika kami melihat pasangan sifat ontogenetik yang diharapkan mewakili fungsi yang sama pada tahap benih dan kecambah, kami mengidentifikasi hubungan yang kuat antara analog ukuran (toleransi stres) (Gambar 6a,b ), tetapi hubungan yang lebih lemah antara analog suhu (Gambar 6c,d ) dan hubungan yang lebih kecil antara analog laju dan cahaya (Gambar 6e,f , berturut-turut). Jika digabungkan, hal ini menunjukkan bahwa aspek independen dari fungsi pada setiap tahap dapat meningkatkan dimensionalitas fungsional ketika tahap benih dan bibit digabungkan untuk menggambarkan ceruk perekrutan fungsional.
4 DISKUSI
Kami mengeksplorasi bagaimana sifat benih dan bibit bersama-sama membentuk fungsi rekrutmen sebagai sarana untuk mengkarakterisasi sebagian dari berbagai strategi regenerasi yang diteorikan Grubb ( 1977 ). Di antara lusinan spesies restorasi semi-kering, pola korelasi sifat mengidentifikasi kemungkinan dimensi fungsional dalam tahap benih dan bibit, namun juga mengungkapkan bagaimana fungsi dapat dan mungkin tidak dikoordinasikan di seluruh ontogeni awal. Kami mengidentifikasi setidaknya dua dimensi fungsional yang menghubungkan aspek strategi benih dan bibit pada spesies padang rumput beriklim sedang ini; namun, penilaian kami juga menunjukkan bahwa relung fungsional rekrutmen bukanlah serangkaian trade-off linier sederhana, tetapi struktur koordinasi sifat morfo-fisiologis yang berpotensi kompleks dengan berbagai tingkat sinergi fungsional di seluruh tahapan.
4.1 Dimensi fungsional benih
Dalam mendeskripsikan aspek-aspek utama fungsi benih, Saatkamp et al. ( 2019 ) menyarankan setidaknya tiga dimensi yang memisahkan spesies, termasuk pertukaran penyebaran-pembentukan benih, persistensi dalam tanah dan waktu perkecambahan. Kami mengukur sifat-sifat dengan hubungan potensial dengan toleransi stres selama pembentukan (massa benih) dan persistensi dalam tanah (bentuk dan massa benih), tetapi memprioritaskan waktu perkecambahan mengingat hubungan langsungnya dengan kondisi lingkungan yang dialami oleh bibit dalam ekosistem semi-kering musiman. Kami mengamati sedikit hubungan di antara sifat-sifat yang menggambarkan laju perkecambahan, penundaan (yaitu dormansi benih), respons cahaya dan suhu, yang menunjukkan bahwa waktu perkecambahan mungkin rumit, yang mencerminkan pengaruh gabungan dari beberapa aspek independen dari fungsi perkecambahan dan respons lingkungan (misalnya Chen et al., 2024 ; Dalziell et al., 2022 ). Studi lain juga menemukan independensi di antara faktor-faktor pendorong waktu perkecambahan (misalnya laju perkecambahan dan kebutuhan cahaya, Hoyle et al., 2015 ) dan selanjutnya menyarankan bahwa respons terhadap kondisi variabel, seperti suhu yang berfluktuasi atau kejadian hujan variabel (tidak dipertimbangkan dalam studi ini) juga dapat membedakan strategi (Torres-Martínez et al., 2017 ; Xu & Du, 2023 ). Namun, sinergi di antara sifat-sifat waktu perkecambahan juga telah dilaporkan pada skala yang lebih luas, ketika iklim makro dapat membatasi kemungkinan strategi, yang menekankan perlunya mengevaluasi pola di seluruh sistem dan skala yang berbeda. Misalnya, tanaman yang terbatas di pegunungan Alpen memiliki persyaratan stratifikasi dingin dan suhu perkecambahan hangat relatif terhadap spesies dengan distribusi yang lebih luas (Fernandez-Pascual et al., 2021 ).
Meskipun korelasi umumnya lemah antara massa benih, bentuk dan sifat perkecambahan, kami mengidentifikasi satu dimensi terkoordinasi yang menunjukkan bahwa spesies berbiji besar mampu berkecambah pada suhu yang lebih dingin daripada spesies berbiji lebih kecil (Lampiran S1 : Gambar A7A). Sinergi serupa ditemukan dalam sistem musiman beriklim sedang dingin lainnya (misalnya Alpen atau Mediterania; Arène et al., 2017 ; Carta et al., 2022 ; Fernandez-Pascual et al., 2021 ), yang menunjukkan bahwa cadangan benih yang lebih besar dapat mendukung pembentukan dan memungkinkan perkecambahan lebih awal dalam kondisi basah yang lebih dingin yang dapat dikaitkan dengan bahaya lingkungan (misalnya beku). Meskipun kami tidak menangkap variasi dalam sifat penyebaran benih, mereka juga dapat menunjukkan hubungan penting dengan strategi pembentukan, yang memengaruhi kondisi yang dialami oleh benih di ruang angkasa dan berpotensi mengurangi kebutuhan untuk menoleransi kondisi yang keras (Saatkamp et al., 2019 ).
Tanpa mengintegrasikan filogeni, bentuk biji yang lebih bulat juga terkait lemah dengan atribut yang dapat memperlambat perkecambahan di tanah, termasuk sensitivitas cahaya biji, dormansi, dan laju perkecambahan yang lebih lambat, yang berpotensi mendukung gagasan bahwa morfologi biji berkontribusi pada persistensi dalam beberapa derajat (Long et al., 2015 ). Hilangnya hubungan ini dengan pendekatan filogenetik dan dalam kelompok taksonomi (misalnya bentuk biji dan dormansi, Lampiran S1 : Gambar A7B) dapat menunjukkan peran nenek moyang bersama yang mendorong kovariasi sifat. Namun, penelitian lain mendokumentasikan hubungan persisten antara bentuk biji dan waktu perkecambahan sambil memperhitungkan variasi filogenetik (misalnya Barak et al., 2018 ). Kovariasi yang diamati di antara nilai sifat mentah di sini masih bisa bersifat adaptif jika perbedaan besar dalam bentuk biji (misalnya biji lebih bulat, yang mengakibatkan penguburan dan perlindungan) berevolusi bersamaan dengan kecenderungan dormansi yang lebih kuat (misalnya skarifikasi, yang menyebarkan waktu perkecambahan) pada saat yang sama ketika famili tumbuhan utama dengan sifat-sifat ini (misalnya Fabaceae) menyimpang dari kelompok lain (Westoby et al., 2023 ).
4.2 Dimensi fungsional bibit
Pada tanaman dewasa, sifat ukuran tidak bergantung pada sifat yang terkait dengan perolehan cahaya daun (SLA) dan perolehan sumber daya di bawah tanah (diameter akar; Weigelt et al., 2021 ), yang dapat mengarah pada berbagai strategi di atas tanah (Diaz et al., 2016 ). Sebaliknya, kami menemukan bahwa bibit yang lebih besar secara konsisten memiliki akar yang lebih tebal dan SLA yang lebih rendah setelah perkecambahan (misalnya Larson et al., 2020 ; Maranon & Grubb, 1993 ), yang menunjukkan pengurangan dimensionalitas dengan satu trade-off yang memisahkan bibit yang berkecambah—tumbuh besar (bibit besar) atau tumbuh efisien (SLA tinggi, akar tipis). Sifat morfologi ini menunjukkan konservasi filogenetik yang kuat, dan ada kemungkinan bahwa evolusi dan kendala fisik menyatukan ukuran, cahaya, dan fungsi di bawah tanah tepat setelah perkecambahan, sebelum pengaruh heterogenitas lingkungan, pergeseran ontogenetik, atau faktor lain pada nilai sifat (Laughlin et al., 2017 ).
Ciri bibit yang mencerminkan laju pertumbuhan (RER) dan alokasi untuk perolehan sumber daya di bawah tanah (RMR) ditangkap tepat di luar dimensi ukuran ini, tetapi korelasi sedang antara RER dan biomassa bibit menunjukkan kecenderungan bibit besar untuk menumbuhkan akar dengan cepat. Akibatnya, sementara bibit kecil dikaitkan dengan pertumbuhan relatif yang lebih cepat (Butterfield & Briggs, 2011 ; Maranon & Grubb, 1993 ), ukuran besar dapat berkorelasi dengan pertumbuhan absolut yang lebih cepat setelah perkecambahan (tetapi berkurang dengan perkembangan, Garbowski et al., 2021 ). Namun, karena RER dan RMR dapat merespons lingkungan dengan cepat (Nguyen et al., 2017 ), perbedaan fisiologis juga dapat memengaruhi variasi interspesifik dan memisahkan akar dari morfologi bibit. Sejalan dengan ini, kami mengamati variasi luas dalam suhu minimum untuk pemanjangan akar di seluruh spesies (0°C–8°C) yang tidak terkait dengan morfologi dan RER pada suhu yang lebih hangat. Respons suhu bibit hampir tidak pernah diteliti dalam sistem alami tetapi perlu mendapat perhatian sebagai pendorong potensial fenologi dan keberhasilan dalam mengubah iklim (Mummey et al., 2016 ).
4.3 Koordinasi ontogenetik
Kami menemukan bukti campuran untuk koordinasi ontogenetik antara sifat-sifat yang mewakili fungsi serupa dalam biji dan bibit (toleransi stres terkait ukuran, laju perkecambahan atau pertumbuhan, suhu dan respons cahaya). Kami mengamati hubungan terkuat antara sifat-sifat ukuran (Gambar 6a,b ), yang menunjukkan bahwa kendala fisik pada morfologi dapat memberikan dasar yang kuat untuk koordinasi ontogenetik di seluruh tahap rekrutmen awal, dengan implikasi penting untuk kemunculan dan pembentukan bibit dalam konteks stres abiotik (misalnya Gardarin et al., 2016 ; Lamont & Groom, 2013 ). Aspek fisiologis fungsi lebih kompleks: kami menemukan bukti campuran untuk hubungan ontogenetik antara sifat-sifat yang terkait dengan respons cahaya dan suhu, dan tidak ada hubungan antara sifat-sifat terkait laju pada tahap biji dan bibit.
Kami menduga benih yang membutuhkan cahaya untuk perkecambahan memiliki bibit dengan daun lebih tipis (SLA lebih tinggi) untuk memanfaatkan ketersediaan cahaya dengan cepat (Donohue et al., 2010 ). Meskipun kami tidak menemukan korelasi yang jelas, kami mengamati sedikit variasi dalam sensitivitas cahaya benih (seringkali perbedaan ±10% dalam perkecambahan, Gambar 6f ), yang mungkin juga menjelaskan kurangnya korelasinya dengan sifat-sifat lainnya. Tidak ada penelitian sejauh pengetahuan kami yang mengaitkan kebutuhan cahaya benih dengan SLA atau morfologi bibit, tetapi yang lain telah menemukan hubungan dengan morfologi benih yang juga tidak ada dalam penelitian kami (Fernandez-Pascual et al., 2021 ). Kami juga memperkirakan benih yang berkecambah dingin mampu tumbuh pada suhu yang lebih dingin tetapi menemukan bahwa sifat khusus itu penting: suhu minimum RER berasosiasi lemah dengan suhu minimum perkecambahan tetapi tidak dengan indeks suhu benih (Gambar 6c,d ), sedangkan yang terakhir memiliki asosiasi yang lebih kuat (negatif) dengan RER pada 18°C (Lampiran S1 : Gambar A7). Ini menunjukkan koordinasi yang lemah di antara ambang batas suhu terdingin untuk perkembangan (misalnya Gardarin et al., 2016 ), tetapi dasar yang lebih kuat untuk mengharapkan bahwa spesies yang berkecambah dingin memiliki pertumbuhan yang lebih cepat setelah suhu menghangat di padang rumput musiman semi-kering ini. Sebaliknya, kami tidak menemukan hubungan antara metrik tingkat (tingkat perkecambahan dan RER), meskipun yang lain telah mendokumentasikan hubungan berdasarkan metrik pertumbuhan relatif (misalnya Shipley & Parent, 1991 ). Secara umum, kami mengamati sedikit korelasi dengan tingkat perkecambahan, yang dipengaruhi oleh dormansi, cahaya dan suhu dan mungkin kurang relevan setelah faktor-faktor ini diperhitungkan (seperti dalam metrik tingkat perkecambahan kami), atau relatif terhadap metrik waktu perkecambahan lainnya seperti sinkronisasi, penyebaran dan respons terhadap kondisi variabel (Lozano-Isla et al., 2019 ; Xu & Du, 2023 ).
4.4 Koordinasi fungsional
Ketika kami mempertimbangkan sifat-sifat yang mewakili fungsi-fungsi yang berbeda di seluruh tahap ontogenetik, kami menemukan beberapa sinergi dan trade-off yang menunjukkan bahwa waktu perkecambahan dapat dikaitkan dengan fungsi bibit dengan cara-cara yang tidak terduga tetapi bermakna. Hubungan antara lingkungan perkecambahan dan strategi pembibitan yang sesuai ini menawarkan wawasan tentang bagaimana berbagai jendela rekrutmen temporal dapat didukung di antara spesies yang hidup berdampingan dalam ekosistem musiman, semi-kering. Mereka juga menunjukkan bahwa tahap perkecambahan dan pembibitan awal mungkin lebih terkait erat, seperti yang diharapkan dalam ekosistem dengan bahaya rekrutmen abiotik yang sering terjadi (Donohue et al., 2010 ), dibandingkan dengan ikatan yang lebih lemah yang kadang-kadang diamati antara strategi perkecambahan dan sifat-sifat dewasa (Hoyle et al., 2015 ).
Dimensi fungsional pertama (PC1) mengungkapkan bahwa spesies dengan biji dan bibit yang lebih besar, daun dan akar yang lebih tebal, dan pemanjangan akar yang lebih cepat cenderung berkecambah pada suhu yang lebih dingin. Bersama-sama, ini menunjukkan bahwa perkecambahan awal dan dingin mungkin menguntungkan ketika cadangan benih yang lebih besar mendorong bibit yang kuat dan tumbuh cepat dengan sifat-sifat yang dapat dikaitkan dengan toleransi beku pada bibit (Bucher & Rosbakh, 2021 ). Dimensi fungsional kedua (PC2) menunjukkan bahwa spesies dengan tingkat dormansi benih yang lebih rendah cenderung berinvestasi pada akar, dengan alokasi biomassa yang lebih besar (RMR) dan RER yang lebih cepat. Dormansi benih adalah mekanisme untuk menghindari atau menyebarkan risiko karena kondisi lingkungan yang keras melalui penundaan perkecambahan (ten Brink et al., 2020 ), sementara investasi akar menyediakan mekanisme untuk bertahan hidup dalam kondisi yang keras dengan meningkatkan akses ke, atau toleransi terhadap, sumber daya yang terbatas pada tahap pembibitan (Garbowski et al., 2020 ). Secara keseluruhan, dimensi ini dapat mewakili trade-off ontogenetik untuk menghadapi lingkungan yang keras dengan menghindarinya pada tahap benih (melalui dormansi) atau bertahan hidup melaluinya pada tahap pembibitan (melalui penanaman akar), meskipun sedikit penelitian sejauh pengetahuan kami telah mengeksplorasi kaitan ini. Sementara hubungan ini kuat terhadap filogeni, hubungan ini juga tampak lebih lemah dalam beberapa kelompok taksonomi (misalnya Asteraceae ) dan berpotensi lebih kuat sehubungan dengan mekanisme dormansi tertentu (misalnya yang melibatkan stratifikasi, Lampiran S1 : Gambar A7).
4.5 Dimensi ceruk perekrutan fungsional
Dalam tahap benih dan bibit, empat atau tiga dimensi diperlukan untuk menjelaskan variasi di tujuh atau enam sifat, masing-masing. Beberapa dimensi ini selaras untuk menstabilkan jumlah dimensi yang disarankan untuk menangkap variasi fungsional di kedua tahap rekrutmen ( n = 4). Dimensi pertama (sifat PC1,7), yang menyatukan sifat toleransi stres terkait ukuran dengan kapasitas untuk berkecambah dalam kondisi dingin, merupakan pendorong utama stabilitas ini. Mengingat kekuatan dan sifat mekanistik hubungan sifat yang mendasarinya, dan konsistensinya di seluruh famili tanaman utama (misalnya massa benih dan indeks suhu benih, Lampiran S1 : Gambar A7A), asosiasi serupa mungkin diharapkan dalam sistem semi-kering lainnya di mana rekrutmen dipengaruhi oleh dinamika musim dingin. Dalam dimensi ini, bagaimanapun, ikatan yang lebih kuat dan lebih konsisten terjadi di antara sifat-sifat dengan dasar morfologi (yaitu ukuran benih dan bibit, diameter akar, SLA) relatif terhadap sifat fisiologis yang mencerminkan laju pertumbuhan (RER) dan respons lingkungan (metrik suhu benih). Di luar dimensi ini, independensi beberapa sifat fisiologis lainnya menyebabkan ceruk rekrutmen fungsional yang tidak seimbang, dengan dimensi ketiga dan keempat menangkap sifat-sifat individual dengan variasi yang tidak dapat dijelaskan (suhu minimum RER) atau sedikit variasi di antara spesies yang diteliti (indeks cahaya benih). Dua sifat lainnya tidak terwakili dengan baik oleh PCA, dengan peran fungsional yang kurang jelas setelah memperhitungkan filogeni (bentuk benih) atau pendorong perkecambahan lainnya (laju perkecambahan, diperkirakan setelah persyaratan cahaya dan dormansi terpenuhi).
Pola-pola ini menunjukkan bahwa dimensionalitas ceruk rekrutmen fungsional bisa lebih tinggi jika rentang sifat yang lebih lengkap yang mencerminkan fisiologi dan respons lingkungan disertakan dalam penilaian variasi fungsional. Namun, sifat-sifat ini kurang dipelajari, dan masih banyak yang harus dipelajari tentang sifat fisiologis mana yang paling baik menangkap respons fungsional (Jimenez-Alfaro et al., 2016 ), dan bagaimana ini bervariasi di berbagai ekosistem. Hal ini berlaku untuk tahap dewasa juga, di mana sintesis yang mengidentifikasi empat hingga enam dimensi telah memusatkan sifat morfologi, dengan lebih sedikit representasi dan pemahaman di sekitar sifat yang menangkap perbedaan berkelanjutan dalam fisiologi (Laughlin, 2014 ; Weigelt et al., 2021 ). Karena kinerja benih dan bibit sangat bergantung pada respons jangka pendek dan skala kecil terhadap lingkungan, sifat fisiologis yang memengaruhi respons ini layak mendapat perhatian lebih besar dalam organisasi teoritis dan karakterisasi empiris ceruk rekrutmen fungsional. Meskipun terdapat potensi peningkatan dimensionalitas, hasil kami sejalan dengan hasil penelitian lain yang menunjukkan bahwa koordinasi ontogenetik dan fungsional dapat menyederhanakan beberapa aspek morfo-fisiologis dari strategi perekrutan (misalnya Funk et al., 2024 ).
4.6 Melangkah maju
Mengukur sifat-sifat awal dapat membantu menjelaskan struktur dan implikasi variasi dalam ceruk perekrutan, tetapi sangat penting untuk mensintesis pola di seluruh tahap ontogenetik. Mengikat pendorong spesifik waktu perkecambahan (misalnya suhu atau dormansi) ke fungsi bibit (misalnya ukuran atau investasi akar) memberikan pandangan yang lebih holistik tentang stresor lingkungan yang kemungkinan besar dialami dan ditoleransi oleh bibit, masing-masing, dan kapan ketidakcocokan dapat terjadi – misalnya, jika waktu perkecambahan bergeser dalam iklim yang berubah (misalnya Mattana et al., 2023 ). Lebih jauh lagi, beberapa aspek fungsi mungkin diperlukan untuk memahami perbedaan rekrutmen yang memungkinkan koeksistensi atau pengecualian di antara spesies. Misalnya, kami menyertakan dua spesies rumput eksotis yang sering bersaing dengan penduduk asli yang diunggulkan selama restorasi. Bersama-sama, atribut benih dan bibit mereka menunjukkan jalur terkoordinasi untuk keberhasilan kompetitif awal dan kuat di gurun dingin, termasuk perkecambahan dalam suhu dingin dan dormansi benih rendah yang dipasangkan dengan investasi dalam pemanjangan akar dan biomassa (Lampiran S1 : Gambar A7B,C).
Pada saat yang sama, beberapa keterbatasan studi kami merupakan tantangan yang harus diatasi. Kumpulan spesies kami dibatasi oleh keragaman taksonomi spesies restorasi umum dalam sistem ini, dan sementara kami memprioritaskan kuantifikasi variasi antar spesies, variasi sifat benih dan bibit juga dipengaruhi oleh adaptasi lokal, efek maternal dan plastisitas (misalnya Fernández-Pascual et al., 2019 ; Vasey et al., 2023 ). Lebih jauh lagi, mengidentifikasi sifat mana yang memberikan indikator perbedaan paling jelas di antara spesies merupakan tantangan yang berkelanjutan (Winkler et al., 2024 ). Untuk fungsi yang ditargetkan, beberapa sifat dapat dipilih atau dievaluasi dengan cara yang berbeda (misalnya untuk waktu perkecambahan atau respons cahaya, Lozano-Isla et al., 2019 ; Zhang et al., 2022 ), sementara metrik tunggal mungkin tidak menangkap kompleksitas penuh respons lingkungan (misalnya Sánchez et al., 2014 ). Kami memilih sifat-sifat untuk mewakili fungsi-fungsi utama dalam sistem studi kami (respons cahaya, suhu) tetapi kemungkinan besar melewatkan sifat-sifat lain yang dapat memengaruhi dinamika rekrutmen dan dimensionalitas relung (misalnya respons benih terhadap air, penyebaran, dan respons terhadap kondisi variabel). Ke depannya, penyelesaian ketidakpastian tentang relevansi sifat akan memerlukan upaya empiris yang tidak hanya menangkap rentang penuh variasi sifat dalam dan di antara spesies, tetapi juga secara bersamaan menguji peran fungsional sifat-sifat rekrutmen ini dalam lingkungan nyata.
KONTRIBUSI PENULIS
Julie Larson dan Stella Copeland menggagas gagasan dan merancang metodologi; Julie Larson dan Dylan Neuhaus mengumpulkan data; Julie Larson menganalisis data dan memimpin penulisan naskah. Semua penulis memberikan kontribusi kritis terhadap draf dan memberikan persetujuan akhir untuk publikasi. Pernyataan tentang penyertaan: Julie Larson dan Stella Copeland meminta masukan tentang pemilihan spesies dan sifat selama proses desain studi dari organisasi dan lembaga federal yang terlibat dalam implementasi dan penelitian restorasi skala besar di AS bagian barat, termasuk karyawan Biro Pengelolaan Lahan, The Nature Conservancy, Survei Geologi AS, Dinas Perikanan dan Satwa Liar AS, dan Dinas Kehutanan AS. Julie Laron membagikan hasil studi dalam presentasi kepada pengelola lahan dan pembuat keputusan.
UCAPAN TERIMA KASIH
Kami berterima kasih kepada S. Kulpa, K. Gunnell, L. Svejcar, C. Guetling, B. Butterfield atas penyediaan benih, dan M. Slate atas masukan pada draf awal naskah. Kami berterima kasih kepada L. Ziegenhagen, R. Rios, D. Golembiewski, M. Willoughby, J. Christy, S. Sternick, Z. Vavrek, dan lainnya di Eastern Oregon Agricultural Research Station atas dukungan mereka di laboratorium.
PERNYATAAN KONFLIK KEPENTINGAN
Penulis menyatakan tidak ada konflik kepentingan.