
ABSTRAK
Tujuan
Play Face (PF) dan Full Play Face (FPF) pada kera besar—homolog dengan wajah tersenyum dan tertawa manusia—dianggap sebagai fenomena tunggal. Namun, jika seleksi alam telah melestarikan dua ekspresi, mungkin nilai adaptifnya berbeda.
Bahan dan Metode
Kami mengumpulkan data video tentang interaksi bermain pada dua kelompok gorila dataran rendah ( N = 21; Gorilla gorilla gorilla ) yang ditempatkan di La Vallée des Singes dan ZooParc de Beauval (Prancis). Karena tidak memiliki alat yang dirancang khusus untuk gorila selama penelitian ini, kami menganalisis aktivasi unit aksi wajah melalui chimpFACS dan OpenFace.
Hasil
Kami menemukan bahwa PF dan FPF mengaktifkan unit tindakan yang sebagian berbeda seperti yang terjadi pada simpanse dan PF/FPF manusia. Kami mendeteksi replikasi cepat (Rapid Facial Mimicry [RFM]) dari PF atau FPF yang dikaitkan dengan sesi bermain yang lebih lama. PF yang tidak ditiru dikaitkan dengan peningkatan variabilitas sesi bermain (berbagai jenis pola bermain) yang diukur melalui Indeks Shannon, sedangkan FPF yang tidak ditiru dikaitkan dengan peningkatan asimetri bermain (ketidakseimbangan antara pola menyerang/bertahan) yang diukur melalui Indeks Asimetri Bermain.
Diskusi
Gorila dataran rendah dapat menggunakan PF untuk mengelola sesi yang lebih kompleks dalam hal jenis pola dan FPF—sinyal yang lebih menonjol—untuk mencegah kesalahpahaman saat sesi tidak seimbang. RFM dari kedua ekspresi tersebut dapat mendukung perpanjangan sesi bermain dengan meningkatkan sinkronisasi pemain dan kemungkinan berbagi emosi. Studi kami membuka pintu untuk studi perbandingan lebih lanjut tentang ekspresi main-main pada manusia dan primata lain sebagai cara untuk menyempurnakan kemungkinan komunikasi emosional dan menggambarkan akar evolusi potensial dari komunikasi wajah Hominidae.
1 Pendahuluan
![]()
Ekspresi wajah—gerakan wajah yang terlihat yang terkait dengan perilaku khusus spesies yang memiliki makna komunikatif bagi anggota kelompok (Waller et al. 2020 )—bukanlah ciri khas manusia. Mamalia lain menunjukkan fenomena perilaku serupa dalam hal bentuk dan fungsi, sehingga menunjukkan bahwa ekspresi wajah telah berevolusi jauh sebelum munculnya manusia modern (Waller dan Micheletta 2013 ).
Pada Homo sapiens , senyum dan tawa memegang peranan penting dalam kohesi sosial, dan kemungkinan besar telah berevolusi dalam konteks sosial bermain (Provine 2000 ; Dezecache dan Dunbar 2012 ; Bryant et al. 2016 ) di mana primata manusia dan nonmanusia menunjukkan ciri-ciri anatomi dan fungsional yang sama dalam perilaku wajah (Cordoni dan Palagi 2011 ; Davila-Ross dan Palagi 2022 ). Terdapat berbagai varian senyum dan tawa yang lebih atau kurang tulus pada manusia (Ambadar et al. 2009 ; Mascaró et al. 2021 ). Senyum spontan dan komponen visual dari tawa (selanjutnya disebut wajah tertawa) memiliki aktivasi saraf yang sama (terutama di area motorik suplementer bilateral; Iwase et al. 2002 ) dan fitur morfologi dasar yang melibatkan kontraksi otot-otot wajah seperti zygomaticus mayor (sudut bibir tertarik ke belakang dan ke atas), orbicularis oculis (kaki gagak), dan orbicularis oris (menekan bibir; Parr et al. 2007 ; Caeiro et al. 2013 ). Namun, senyum dan wajah tertawa tidak sepenuhnya tumpang tindih.
Senyum spontan manusia dan wajah tertawa tampaknya homolog dengan ekspresi mulut terbuka rileks yang diamati hampir secara eksklusif selama bermain pada primata non-manusia, yaitu, Wajah Bermain (PF) dan Wajah Bermain Penuh (FPF), masing-masing (van Hooff dan Preuschoft 2003 ; Davila-Ross dan Dezecache 2021 ). PF—yang hampir ada di semua spesies primata (Palagi, Burghardt, et al. 2016 )—biasanya melibatkan paparan gigi bawah, sedangkan selama FPF—yang memiliki distribusi yang lebih tidak merata daripada PF dalam primata (Palagi, Burghardt, et al. 2016 )—juga gigi atas biasanya terekspos karena kontraksi otot levator labii superioris (Davila-Ross et al. 2015 ; Waller et al. 2020 ). Pada simpanse ( Pan troglodytes )—seperti pada manusia—baik PF maupun FPF melibatkan kontraksi zygomaticus mayor (Parr et al. 2007 ; Waller et al. 2015 , 2020 ). Lebih jauh lagi, selama FPF, simpanse, seperti manusia, dapat memperlihatkan bibir yang terbuka, menarik bibir ke belakang dan ke atas, rahang ternganga, bibir atas terangkat, dan terkadang pipi terangkat (Davila-Ross et al. 2015 ). Perlu dicatat bahwa tingkat aktivasi dari berbagai unit aksi otot wajah (AU) yang terlibat dalam kinerja PF dan FPF dapat menunjukkan tingkat variabilitas tertentu, yang menyoroti bahwa kedua ekspresi tersebut mewakili gradien intensitas. Misalnya, pada simpanse di tempat perlindungan, Davila-Ross et al. ( 2015 ) menemukan bahwa 19% PF menunjukkan aktivasi otot-otot yang terkait dengan terangkatnya bibir atas, sementara Parr et al. ( 2007 ) tidak menemukan aktivasi semacam itu pada simpanse Yerkes. Oleh karena itu, bukan hanya paparan gigi atas tetapi juga ukuran bukaan mulut dapat menjelaskan perbedaan antara PF dan FPF (Davila-Ross et al. 2015 ).
Deteksi dan analisis otomatis AU wajah merupakan salah satu cara utama untuk mengidentifikasi dan menganalisis berbagai ekspresi wajah. Melalui perangkat khusus (misalnya, Facial Action Coding System [FACS]: Ekman dan Friesen 1978 ; Ekman et al. 2002 ; OpenFace: Baltrušaitis et al. 2018 ), perubahan dalam kontraksi otot wajah yang terkait dengan ekspresi yang berbeda telah diidentifikasi pada manusia (Gilbert et al. 2021 ; Jeganathan et al. 2022 ), simpanse (Vick et al. 2007 ), kera rhesus (Parr et al. 2010 ), siamang (Waller et al. 2012 ), orangutan (Caeiro et al. 2013 ), anjing (Waller et al. 2013 ), kucing (Caeiro et al. 2017 ), dan kuda (Wathan et al. 2015 ).
Menurut Hipotesis Kompleksitas dan Kontinuitas (Davila-Ross dan Dezecache 2021 ), kesamaan yang kuat dalam morfologi dan konteks menunjukkan adanya kesinambungan evolusi, tanpa perubahan besar, dari hominid non-manusia hingga manusia modern.
Selama interaksi yang menyenangkan, fungsi utama dari wajah tersenyum/tertawa dan PF/FPF adalah untuk memberi sinyal niat perilaku non-agresif dari agen, meskipun dengan tingkatan emosional yang berbeda (Pellis dan Pellis 1996 ; Gervais dan Wilson 2005 ; Cordoni dan Palagi 2011 , 2012 ; Palagi et al. 2022 ). Ini khususnya penting selama bermain perkelahian, suatu bentuk permainan di mana banyak pola perilaku diambil dari konteks agresif, dan kontak fisik antara pemain dapat dipertahankan untuk waktu yang lama (Paquette 1994 ; Palagi, Burghardt, et al. 2016 ; Pellis dan Pellis 2017 ; Cordoni et al. 2023 ). Hipotesis Asimetri Kekuasaan (PAH, Preuschoft dan van Hooff 1997 ) mengasumsikan bahwa spesies primata yang hidup dalam masyarakat despotik yang dicirikan oleh hubungan hierarkis yang sangat asimetris perlu menghasilkan sinyal yang tidak ambigu dan berbeda untuk membedakan permainan (terutama, perkelahian sambil bermain) dari interaksi lain seperti peredaan atau afiliasi. Meskipun PAH awalnya dibingkai untuk menjelaskan variasi antar spesies dalam pola tampilan afiliatif/peredaan/main-main, telah disarankan bahwa itu juga dapat diterapkan dalam spesies yang menunjukkan variasi besar dalam hubungan dominasi antara anggota kelompok (Mehu dan Dunbar 2008 ). Selama interaksi bermain, asimetri antara pemain dalam pertukaran pola perilaku dan pencapaian posisi pemenang dapat terjadi (Bauer dan Smuts 2007 ; Pellis et al. 2010 ; Pellis dan Pellis 2017 ). Misalnya, pada simpanse, peningkatan penggunaan ekspresi bermain dikaitkan dengan peningkatan asimetri bermain dengan tujuan yang mungkin untuk mengurangi kesalahpahaman antara pemain ketika permainan menjadi lebih kompetitif (Cordoni dan Palagi 2011 ).
Pada manusia, senyum dan wajah tertawa dapat menyampaikan sinyal dengan intensitas yang berbeda (Redican 1982 ). Namun, senyum bukan hanya tanda kebahagiaan (Ekman 2003 ). Bergantung pada keadaan, senyum dapat menunjukkan kegugupan, kebutuhan untuk menyenangkan, rasa malu, sikap menyambut, dan makna yang berbeda ini dapat dikaitkan dengan variasi spesifik dalam fitur morfologi dan dinamis senyum (Ambadar et al. 2009 ). Di sisi lain, wajah tertawa tidak secara eksklusif dianggap sebagai ekspresi humor atau kebahagiaan. Memang, wajah tertawa dengan komponen pendengarannya dapat didefinisikan sebagai perilaku sosial yang sejalan dalam membangun dan mengatur ikatan sosial dan dalam mengurangi ketegangan sosial (Scott et al. 2014 ; Wood dan Niedenthal 2018 ; Palagi et al. 2022 ).
Pada primata non-manusia, PF dan FPF sebagian memiliki morfologi dan fungsi yang serupa (misalnya, memperpanjang sesi, menghindari eskalasi menjadi agresi) dan sering kali diringkas menjadi satu fenomena (Pellis dan Pellis 1996 , 1997 ; Palagi et al. 2007 ; Demuru et al. 2015 ). Namun, pada spesies primata yang berbeda (dan semua spesies hominid) evolusi telah mempertahankan sinyal PF dan FPF (Davila-Ross dan Dezecache 2021 ). Lebih jauh lagi, selama bermain baik PF dan FPF juga dapat ditemukan secara terpisah (Waller dan Cherry 2012 ; Palagi, Norscia, et al. 2019 ). Oleh karena itu, dari sudut pandang adaptif, masuk akal untuk berhipotesis bahwa kedua sinyal ini mungkin tidak memiliki fungsi yang sepenuhnya tumpang tindih. Misalnya, dibandingkan dengan PF, FPF ditemukan terutama terkait dengan permainan intensitas tinggi pada gorila (Waller dan Cherry 2012 ). Menurut literatur, FPF khususnya dapat digunakan: (i) dalam masyarakat yang lebih toleran di mana kecil kemungkinan FPF disalahartikan sebagai ancaman (dalam kelompok despotik seperti beberapa spesies makaka—misalnya, makaka rhesus—atau babun, terbukanya gigi atas dapat menyebabkan agresi oleh kelompok dominan; Thierry et al. 1989 ; van Hooff dan Preuschoft 2003 ); (ii) ketika permainan kasar dan memerlukan seringnya tampilan sinyal permainan yang menonjol untuk mengomunikasikan maksud pemain yang tidak agresif (Palagi, Burghardt, et al. 2016 ; Palagi, Cordoni, et al. 2016 ). Dalam hal ini, pada gorila dataran rendah, permainan dapat meningkat menjadi agresi tanpa sinyal permainan yang menonjol (Palagi et al. 2007 ; Palagi, Marchi, et al. 2019 ; Bresciani et al. 2021 ). Memang, gorila dataran rendah ( Gorilla gorilla gorilla ) dapat menggunakan lebih banyak FPF daripada simpanse mungkin dalam kaitannya dengan sesi bermain yang sangat kasar (Palagi, Norscia, et al. 2019 ), meskipun tidak ada penelitian yang secara jelas menunjukkan hal ini.
Baik wajah tersenyum maupun tertawa pada manusia dan PF/FPF pada kera besar dapat bertindak sebagai pelepas stimulus ( sensu Tinbergen 1952 ) dan memunculkan ekspresi wajah yang sama pada subjek lain yang berinteraksi (lihat ulasan Palagi et al. 2020 ). Ketika replikasi ekspresi wajah yang sama terjadi dalam 1 detik sejak emisi stimulus, fenomena ini dikenal sebagai Rapid Facial Mimicry (RFM; Dimberg dan Thunberg 1998 ; Sestito et al. 2013 ). Replikasi wajah tidak disengaja dan otomatis (seperti yang ditunjukkan oleh kecepatan respons < 1 detik) dan terjadi secara signifikan lebih banyak ketika stimulus pertama (yaitu, ekspresi) dirasakan oleh penerima potensial dibandingkan ketika tidak dirasakan. Namun perbedaan antara kondisi persepsi ya dan tidak persepsi (dari stimulus pertama) menunjukkan bahwa fenomena tersebut adalah mimikri dan bukan sinkronisasi sederhana (Palagi et al. 2020 ). RFM tidak melibatkan proses kognitif tingkat lanjut, melainkan proses dasar otomatis dan tak sadar dengan menemukan akarnya dalam penggabungan otomatis persepsi dan tindakan dalam area sensorimotor otak, seperti yang diramalkan oleh Model Tindakan Persepsi yang mungkin melibatkan sistem neuron cermin (Gallese et al., 1996; Ferrari et al., 2003 ; de Waal dan Preston , 2017 ).
RFM hadir selama bermain pada hewan manusia dan nonmanusia yang menunjukkan tampilan mulut terbuka (lihat ulasan Palagi et al. 2020 ). Pada banyak spesies primata dan nonprimata, kemunculan RFM—dibandingkan dengan kehadiran sederhana PF yang tidak direplikasi—dikaitkan dengan durasi interaksi bermain yang lebih lama, sehingga keberhasilannya lebih tinggi (monyet Tonkean, Scopa dan Palagi 2016 ; gelada, Mancini et al. 2013 ; gorila dataran rendah, Bresciani et al. 2021 ; simpanse, Palagi, Norscia, et al. 2019 ; orangutan, Davila-Ross et al. 2008 ; anjing, Palagi et al. 2015 ; meerkat, Palagi, Marchi, et al. 2019 ).
Pada gorila dataran rendah—meskipun dengan pengecualian (Cordoni dkk. 2022 )—permainan hampir tidak dapat dilakukan saat dewasa (Masi dkk. 2009 ; Cordoni dkk. 2018 ). Gorila menunjukkan PF dan FPF (Palagi dkk. 2007 ; Bresciani dkk. 2021 ) dan kemunculan RFM—hanya diverifikasi dengan menggabungkan PF dan FPF—telah ditunjukkan selama sesi permainan sosial mereka (Palagi, Norscia, dkk. 2019 ; Bresciani dkk. 2021 ). Selain itu, gorila memiliki sejumlah besar otot wajah seperti Hominoidea lainnya (misalnya, zygomaticus mayor, orbicularis oculi, levator labii superioris; Diogo et al. 2009 , 2010 ). Oleh karena itu, gorila sangat cocok untuk menyelidiki dasar evolusi perbedaan antara wajah tersenyum dan wajah tertawa. Untuk tujuan ini, kami menyelidiki perbedaan morfologi (yaitu, aktivasi AU otot wajah) dan fungsi antara PF dan FPF serta replikasi cepat (RFM) pada gorila dataran rendah. Berdasarkan kerangka kerja sebelumnya, kami merumuskan prediksi berikut.
1.1 Prediksi 1
Bahasa Indonesia: Jika PF dan FPF adalah ekspresi dasar yang telah dilestarikan dalam perjalanan evolusi ekspresi wajah primata (Waller et al. 2020 ), kami berharap bahwa—dengan menerapkan alat yang digunakan untuk analisis ekspresi pada hominid lain (manusia–OpenFace, simpanse–ChimpFACS)—kami akan mendeteksi dengan kesalahan minimal aktivasi unit otot wajah yang sama yang aktif dalam PF dan FPF manusia dan simpanse ( Prediksi 1a ). Selain itu, karena PF dan FPF seharusnya tidak memiliki morfologi yang sama persis, kami berharap bahwa unit wajah yang aktif selama dua ekspresi tersebut tidak sepenuhnya tumpang tindih ( Prediksi 1b ).
1.2 Prediksi 2
Karena RFM hadir pada gorila (Palagi, Norscia, dkk. 2019 ; Bresciani dkk. 2021 ) dan PF dan FPF dapat terjadi secara terpisah selama bermain, kami berharap menemukan RFM baik PF maupun FPF ( Prediksi 2a ). Lebih jauh, kami berharap durasi sesi bermain mungkin lebih lama dengan adanya RFM baik PF maupun FPF daripada dengan adanya sinyal wajah yang tidak direplikasi atau tidak ada sinyal ( Prediksi 2b ).
1.3 Prediksi 3
Pada gorila, permainan sangat kompetitif dan asimetris, terutama antara jantan muda/remaja (Palagi et al. 2007 ; Cordoni et al. 2018 ). Dalam hal ini, kami berharap bahwa FPF—yang lebih merupakan sinyal menonjol daripada PF (Palagi dan Mancini 2011 ; Palagi, Burghardt, et al. 2016 )—dapat dikaitkan dengan tingkat asimetri permainan yang tinggi untuk menyampaikan pernyataan suasana hati positif yang lebih jelas oleh agen ( Prediksi 3a ). Lebih jauh, kami juga berharap bahwa dalam kasus interaksi yang lebih simetris, sinyal yang kurang jelas seperti PF daripada FPF dapat dikaitkan dengan peningkatan variabilitas permainan (yaitu, berbagai jenis pola perilaku bermain yang dilakukan dalam satu sesi; Prediksi 3b ).
2 Bahan dan Metode
2.1 Pernyataan Etika
Penelitian saat ini murni bersifat observasional dan non-manipulatif; dengan demikian, persetujuan tidak diperlukan oleh komite perawatan hewan institusional penulis.
2.2 Kelompok Belajar
Penelitian ini dilakukan pada dua kelompok keluarga gorila dataran rendah ( Gorilla gorilla gorilla ) yang ditempatkan di La Vallee des Singes (Romagne, Prancis; selanjutnya, VDS) dan ZooParc de Beauval (Saint Aignan sur Cher, Prancis; selanjutnya, BEA).
Kelompok VDS terdiri dari 10 individu (usia rata-rata ± SE: VDS 19,2 ± 5,10 Tabel 1 ). Dua jantan remaja (Mawete dan Djomo) dikebiri. Gorila diberi makan di luar ruangan dengan buah, sayuran, biji, daun, dan batang pohon lima kali sehari selama musim semi/panas dan dua kali sehari mulai bulan September.
| Subjek | Seks | Kelas umur dan tahun lahir | Usia (tahun) a | Kekerabatan | Kelompok | Jumlah jam observasi | Jumlah total sesi bermain | Jumlah total PF | Jumlah total FPF |
|---|---|---|---|---|---|---|---|---|---|
| Yaoundé (YA) | M | Iklan—1983 | 37 | Bapak dari semua SubAd dan Inf | VDS | 92 | 9 | 6 | 14 |
| Hakuna (HA) | F | Iklan—1996 | 24 | Ibu IV | VDS | 80.5 | 24 | 7 | 3 |
| Virunga (VI) | F | Iklan—1970 | 50 | Tidak ada keturunan | VDS | 78 | 2 | 1 | angka 0 |
| Moseka (MO) | F | Iklan—1984 | 35 | MW, DJ, dan KO ibu | VDS | 71.5 | angka 0 | angka 0 | angka 0 |
| Mahmah (MA) | F | Iklan—2002 | 18 | Ibu BA | VDS | 79 | 85 | 48 | 53 |
| Mawete (MW) | M | SubIklan—2011 | 9 | DJ dan KO saudara | VDS | 62.5 | 209 | 70 | 209 |
| Djomo (DJ) | M | SubIklan—2008 | 12 | MW dan KO saudara | VDS | 63 | 43 | 15 | 50 |
| Kouam (KO) | M | Inf—2016 | 4 | MW dan saudara DJ | VDS | 63 | 281 | 70 | 207 |
| Ivindo (IV) | F | Inf—2017 | 3 | — | VDS | 70 | 190 | 32 | 114 |
| Basoko (BA) | M | Inf—2020 | angka 0 | — | VDS | 75 | 86 | 49 | 112 |
| Asato (AS) | M | Iklan—1991 | 30 | Bapak dari semua SubAd dan Inf | BEA | 43.5 | angka 0 | angka 0 | angka 0 |
| Kabinda (KA) | F | Iklan—1982 | 39 | Ibu YA, KO, MY, MA | BEA | 42 | angka 0 | angka 0 | angka 0 |
| Inge (IN) | F | Iklan—1980 | 41 | Ibu KI | BEA | 43 | 2 | 1 | angka 0 |
| Sheila (SH) | F | Iklan—1991 | 30 | Ibu MB, SA | BEA | 42 | angka 0 | angka 0 | angka 0 |
| Mapenzi (MA) | M | SubIklan −2010 | 11 | Saudara kandung MY, KO, YA | BEA | 41 | 8 | 1 | 2 |
| Sawa (SA) | F | SubIklan—2011 | 10 | Saudara kandung MB | BEA | 42 | 3 | angka 0 | angka 0 |
| Mayele (MY) | F | SubIklan—2013 | 8 | Saudara kandung YA, KO, MA | BEA | 41.5 | 2 | 1 | 1 |
| Yamba (bahasa Inggris kuno) | M | SubIklan—2015 | 6 | Saudara kandung MA, MY, KO | BEA | 42.5 | 18 | 2 | 10 |
| Kivano (KI) | M | SubIklan—2016 | 5 | – B | BEA | 42 | 13 | 1 | 2 |
| Mbaku (MB) | M | Inf—2018 | 3 | Saudara kandung SA | BEA | 43 | 28 | 7 | 35 |
| Kovanga (KO) | M | Inf—2019 | 2 | Saudara kandung MA, MY, YA | BEA | 42.5 | 23 | 6 | 19 |
Catatan: Jumlah jam observasi dan jumlah sesi bermain yang melibatkan setiap individu, serta jumlah PF dan FPF yang dilakukan oleh setiap individu dilaporkan dalam tabel. Singkatan: Ad, dewasa (perempuan > 8 tahun; laki-laki > 12 tahun); F, perempuan; Inf, bayi (0–4 tahun); M, laki-laki; SubAd, sub-dewasa (perempuan 6–8 tahun; laki-laki 6–12 tahun). berusia satu tahun pada saat penelitian dilakukan. b Kivano tidak memiliki individu terkait dalam koloninya.
Kelompok BEA terdiri dari 11 individu (usia rata-rata ± SE: 16,82 ± 4,52; Tabel 1 ). Dua jantan muda (Mapenzi dan Yamba) dikebiri. Hewan-hewan tersebut menerima makanan (buah, sayuran, biji-bijian, daun, dan batang pohon) enam kali sehari.
Kelompok VDS dan BEA dikelola dalam kandang serupa yang terdiri dari fasilitas dalam ruangan (VDS 150 m 2 , BEA 200 m 2 ) dan luar ruangan (pulau berhutan yang dikelilingi oleh kanal air; VDS 3400 m 2 , BEA 2000 m 2 ). Kandang diperkaya dengan pohon, liana, batang, jerami, dan platform. Pada siang hari, gorila dapat bergerak bebas antara fasilitas dalam dan luar ruangan dan dapat berinteraksi secara sosial. Di kedua kelompok, gorila punggung perak adalah ayah dari subjek yang belum dewasa, dan semua betina dewasa diobati dengan kontrasepsi oral.
2.3 Pengumpulan Data
Kami mengumpulkan data video di kedua koloni selama periode berikut: Agustus–Oktober 2020/Maret–Juni 2021/Maret–Juni 2022 untuk VDS dan Juni–Oktober 2021 untuk BEA. Untuk perekaman video, digunakan kamera full HD (Panasonic HDC-SD9).
Para pengamat (LP, FG) dilatih oleh GC dalam pengenalan hewan dan penerapan prosedur metodologis (yaitu, metode pemindaian dan pengambilan sampel semua kejadian; lihat di bawah). Selain itu, sebelum memulai analisis video secara independen, para pengamat menerima pelatihan dari GC dalam mengidentifikasi pola perilaku (lihat Tabel 2 ) dan membedakan antara PF dan FPF. Sepuluh persen dari sesi permainan yang direkam (sekitar 100 sesi) dianalisis—baik dalam gerakan lambat atau bingkai demi bingkai menggunakan perangkat lunak gratis Avidemux 2.7.8—oleh kedua pengamat secara bersamaan dan terpisah. Kesepakatan mereka dalam mengidentifikasi pola perilaku dan membedakan antara PF dan FPF dinilai menggunakan kappa Cohen, yang menunjukkan proporsi kesepakatan di luar apa yang diharapkan secara kebetulan. Keandalan antar pengamat antara pembuat kode video dihitung menggunakan fungsi R “cohen.kappa” dan pustaka “irr” dan “psych” (R versi 3.5.3). Pelatihan diakhiri ketika reliabilitas antar pengamat mencapai nilai kappa Cohen sebesar 0,80 untuk PF dan FPF dan dari 0,60 hingga 0,94 untuk pola main-main yang direkam dalam penelitian ini (lihat Tabel 2 ).
| Pola bermain | Definisi |
|---|---|
| Mencoba bermain menggigit | Gorila tersebut mencoba menutup mulutnya pada tubuh pasangannya namun tidak berhasil |
| Intip-intip ajah | Gorila bersembunyi dan tiba-tiba muncul dari tempat perlindungan |
| Bermain menggigit | Gorila menutup mulutnya pada tubuh pasangannya dengan cara yang tidak berbahaya |
| Mainkan serangan mendadak | Gorila melompat dengan keempat anggota tubuhnya ke teman bermainnya, biasanya dalam posisi berkaki empat dan memantul atau tetap di sana. |
| Bermain kejar- kejaran | Gorila berlari di belakang teman bermainnya (permainan sosial) dengan sering mengubah arahnya |
| Bermain memanjat atau berdiri di atas yang lain | Gorila memanjat atau berdiri di tubuh teman bermainnya tanpa mempedulikan posisi teman bermainnya (duduk, berbaring, atau berdiri) |
| Mainkan drag a | Gorila menyeret teman bermainnya dengan mengambilnya dari dahan |
| Mainkan penutup mata a | Gorila menutupi mata teman bermainnya |
| Mainkan lompat a,b , a,b | Gorila melompat pelan-pelan sendiri (dalam hal ini polanya dianggap netral) atau pada teman bermainnya (dalam hal ini polanya dianggap ofensif) hanya dengan kaki yang umumnya dalam posisi bipedal. Lompatan bermain umumnya kecil, sebagian besar diam, dengan sedikit atau tidak ada gerakan maju. |
| Mainkan tendangan a | Gorila dengan lembut menggunakan kakinya untuk memukul teman bermainnya |
| Mainkan tarik a | Gorila menggerakkan teman bermainnya ke arahnya dengan tangan dan kakinya saat bermain |
| Mainkan dorong a | Gorila memindahkan teman bermainnya menjauh darinya dengan tangan dan kakinya |
| Mainkan ambil kembali | Gorila menghalangi teman bermainnya dengan tangannya untuk mencegahnya terbang. Hal ini berbeda dengan tarikan saat bermain yang biasanya dilakukan dengan kedua kaki dan tangan saat bermain. |
| Mainkan tampar a | Gorila menggunakan tangannya yang terbuka untuk memukul bagian tubuh teman bermainnya |
| Mainkan stempel a | Gorila memukul tanah atau teman bermainnya dengan kakinya secara berulang-ulang |
| Bermain tarik tambang | Gorila berusaha menangkap sebuah objek dan menariknya ke arah mereka. |
| Kasar dan berantakan | Gorila bermain dalam kontak fisik yang erat dan terus-menerus dengan menggunakan banyak pola yang dijelaskan dalam tabel ini (misalnya, menggigit, menendang, menampar, menghentakkan kaki). |
| Mainkan Flee c | Gorila berlari menjauh dari pasangannya yang mengejarnya |
| Bermain tempat berteduh c | Gorila melindungi dirinya dari tamparan, gigitan, dll. dari teman bermainnya dengan meletakkan lengannya di atas kepala. |
| Mainkan menggeliat c | Gorila itu menggeliat untuk melepaskan diri dari cengkeraman teman bermainnya |
| Permainan akrobatik b | Gorila berayun sambil menggantung/melompat dari penyangga dan melakukan salto/putaran dalam cara menyendiri atau berkelompok |
| Pesawat terbang b | Gorila yang lebih tua memegang teman bermain yang lebih kecil dengan tangan/kaki di atas kepalanya sambil berbaring di tanah |
| Berputar b | Gorila melakukan salto/putaran pada dirinya sendiri atau bergantung pada tali. Putaran dapat menjadi bagian dari permainan akrobatik |
| Mainkan bawa b | Gorila menggendong teman bermainnya di bagian punggung atau perut (ciri khas pengasuhan anak). Subjek yang digendong berbaring di punggung pembawa atau ia menyentuh perut bagian bawah pembawa dalam semacam pelukan. Pembawa bergerak. |
| Mainkan “beri aku lima” b | Dua gorila berinteraksi tatap muka dan saling menepuk telapak tangan |
| Mainkan ambil b | Gorila dengan lembut memijat teman bermainnya yang memegangnya erat-erat |
| Manipulasi permainan b | Gorila mengambil dan menjelajahi suatu objek tanpa menggunakannya untuk tujuan tertentu |
| Mainkan jalan bulan b | Gorila berjalan mundur, biasanya matanya tetap fokus pada teman bermainnya |
| Mainkan pat b | Gorila tersebut berulang kali dan dengan lembut menyentuh tubuh pasangannya dengan telapak tangannya |
| Mainkan permainan naik gendong b | Gorila diletakkan dengan satu kaki di setiap sisi punggung teman bermainnya dan tubuhnya tegak. Posisi ini tampak seperti penunggang kuda. Pengangkut dapat bergerak atau tidak. |
| Mainkan gulungan b | Gorila memutar tubuhnya dari sisi ke sisi saat berbaring |
| Mainkan goyangkan tali b | Gorila menggerakkan tali yang menjadi tempat teman bermainnya dengan kuat |
| Mainkan slide ke bawah b | Gorila meluncur turun dari bukit, pohon, batu atau peralatan lainnya |
| Mainkan putar balik b | Gorila berlari/berjalan sendiri di sekitar suatu objek tanpa mengubah arahnya |
| Bermain jalan-jalan b | Gorila mengikuti teman bermainnya atau maju mundur |
| jungkir balik b | Gorila membalikkan tubuhnya di atas tanah atau di atas penyangga vertikal secara menyendiri atau berkelompok |
| Menggelitik b | Gorila menggelitik bagian tubuh pasangannya dengan tangan/kakinya |
| Undangan bermain | Gorila melakukan satu atau lebih pola perilaku main-main untuk mengajak rekannya bermain |
| Wajah bermain penuh | Gorila membuka mulutnya dengan gigi atas dan bawah terlihat |
| Mainkan wajah | Gorila membuka mulutnya hanya dengan gigi bawah yang terlihat |
Pola ofensif. b Pola netral. c Pola pertahanan.
Dengan pemindaian pengambilan sampel hewan (Altmann 1974 ) kami mengumpulkan pada interval 10 menit keseluruhan aktivitas harian kelompok dengan mencatat perilaku soliter (bergerak, beristirahat, mencari makan, makan) dan sosial (bermain, perawatan diri, kontak tubuh, agresi, kedekatan). Dengan menggunakan metode ini, kami mengumpulkan total 254 jam pengamatan. Secara khusus, total 60 dari 254 jam dihabiskan oleh hewan untuk bermain (60 jam bermain sesuai dengan 360 pemindaian). Kami juga menggunakan metode pengambilan sampel semua kejadian (Altmann 1974 ) untuk mengumpulkan data tentang interaksi sosial yang menyenangkan: (i) identitas pemain, (ii) pola bermain dalam urutan berurutan (Tabel 2 ), (iii) ekspresi bermain (PF dan FPF) dan durasinya, dan (iv) durasi sesi bermain. Kami membuat lembar Excel di mana, untuk setiap sesi bermain, kami melaporkan semua pola bermain yang diidentifikasi (termasuk PF dan FPF) dalam urutan kejadian berurutan sebagaimana ditentukan oleh analisis video. Penting untuk dicatat bahwa pemindaian dilakukan saat semua pengambilan sampel kejadian berlangsung (total 254 jam dari semua kejadian). Kami menganalisis total 1026 sesi perkelahian bermain (jumlah total sesi bermain yang melibatkan setiap gorila dilaporkan dalam Tabel 1 ). Semua sesi melibatkan kontak fisik antara pemain dan, oleh karena itu, tidak berbeda dalam tingkat intensitas dan kategori permainan. Kami mencatat total 317 PF dan 831 FPF (jumlah total PF dan FPF yang dilakukan oleh setiap gorila selama interaksi bermain mereka dilaporkan dalam Tabel 1 ).
2.4 Identifikasi Unit Wajah—Sistem FACS dan OpenFace
Bahasa Indonesia: Selama studi saat ini, dengan tidak adanya alat yang secara spesifik dirancang untuk mendeteksi aktivasi AU otot wajah pada gorila (GorillaFACS baru-baru ini diimplementasikan oleh Correia-Caeiro et al. 2025 ), kami menggunakan FACS yang diadaptasi untuk simpanse (ChimpFACS; Vick et al. 2007 ) dan OpenFace 2.0 (Ambadar et al. 2009 ; Baltrušaitis et al. 2018 ) untuk manusia untuk mengidentifikasi AU otot wajah spesifik mana yang diaktifkan selama kinerja PF dan FPF oleh gorila. Dengan menggunakan FACS, adalah mungkin untuk membandingkan perilaku wajah secara independen dari variabilitas morfologi wajah di antara individu (misalnya, struktur tulang, timbunan lemak; Waller et al. 2007 , 2008 ). FACS mengidentifikasi kontraksi (biner: 0 = tidak ada kontraksi, 1 = ya kontraksi) dari 33 AU wajah dan sering mengidentifikasi kontraksi sekelompok otot, bukan satu otot.
OpenFace 2.0 dapat mengenali ekspresi wajah melalui pendeteksian aktivasi AU wajah (Amos et al. 2016 ). Ia juga dapat memperkirakan intensitas 17 AU (1, 2, 4, 5, 6, 7, 9, 10, 12, 14, 15, 17, 20, 23, 25, 26, dan 45; Baltrušaitis et al. 2018 ). Karena OpenFace menggunakan detektor wajah berbasis Jaringan Syaraf Tiruan Konvolusional baru dan algoritma deteksi landmark wajah yang dioptimalkan, maka AU juga dapat diidentifikasi saat wajah tidak menghadap ke depan atau/dan dalam kondisi pencahayaan rendah. Perangkat lunak mengekstraksi karakteristik wajah dengan menggunakan histogram gradien berorientasi dan mengurangi dimensionalitas dengan menggunakan PCA. Ekspresi netral khusus subjek diekstraksi dengan menghitung nilai median deskriptor wajah dalam urutan video, dengan asumsi bahwa sebagian besar bingkai berisi ekspresi netral. Wajah median yang diekstraksi dikurangi dari deskriptor fitur, yang menghasilkan fitur yang dinormalkan. Vektor fitur yang dinormalkan menggambarkan perubahan dinamis dari ekspresi netral. Kerangka kerja pengenalan AU menggunakan mesin vektor pendukung kernel linear untuk deteksi kejadian AU dan regresi vektor pendukung untuk estimasi intensitas AU (Baltrušaitis et al. 2018 ).
Kami menerapkan FACS dan OpenFace untuk mengevaluasi aktivasi AU dari PF (untuk delapan gorila terpilih) dan FPF (untuk sembilan gorila terpilih; lihat Gambar 1 dan Video S1A,B ). Untuk setiap individu terpilih, setidaknya satu PF dan satu FPF dianalisis (lihat Tabel S2 ). Karena FACS dan OpenFace tidak dirancang untuk gorila, kami memilih ekspresi wajah (PF dan FPF) yang dapat terlihat sangat jelas pada video dan sebagian besar frontal untuk penilaian awal. Penting untuk dicatat bahwa tujuan utama dari bagian penelitian ini adalah untuk memverifikasi bahwa PF dan FPF berbeda, bukan untuk memberikan indikasi definitif dan halus dari semua AU yang diaktifkan, terutama karena itu akan memerlukan GorillaFACS, yang belum tersedia pada saat penelitian kami (Correia-Caeiro et al. 2025 ). MB mengodekan ekspresi ini dengan FACS (manual untuk pengodean digunakan) dan OpenFace dan mengonfirmasi jenis PF yang ditentukan oleh pengamat (LP dan FG) sebelum pengodean dengan perangkat lunak. Kemudian, IN dan GC mengodekan ulang PF dan FPF gorila yang dipilih oleh FACS dan OpenFace. Keandalan antar-pengode mencapai nilai kappa Cohen yang sama dengan 0,81 (17 PF dan 15 FPF dinilai).

2.5 Definisi Operasional
2.5.1 Sesi Bermain
Sesi bermain dimulai ketika satu individu mengarahkan pola perilaku main-main (Tabel 2 ) ke arah sesama spesies dan berakhir ketika salah satu pemain atau individu ketiga yang tidak terlibat mengganggu interaksi (Cordoni et al. 2021 , 2022 ). Durasi rata-rata (dalam detik) sesi bermain adalah 42,80 ± 2,10 SE, dengan nilai minimum 5,0 detik dan nilai maksimum 405,0 detik. Kami membedakan perilaku menjadi ofensif ( O —perilaku menyerang terhadap pasangan), defensif ( D —perilaku menghindari/melindungi terhadap serangan oleh pasangan), dan netral ( N —bukan perilaku ofensif maupun defensif; lihat Tabel 2 ; Cordoni et al. 2021 , 2022 , 2023 ).
2.5.2 Durasi PF dan FPF
Durasi semua PF dan FPF yang diamati dalam video yang dikumpulkan dihitung dengan analisis video bingkai demi bingkai yang dimulai dari bingkai pertama saat bibir gorila dipisahkan hingga bingkai pertama saat bibir ditutup lagi (Bertini et al. 2021 ; Bresciani et al. 2021 ). Keandalan antar pengamat untuk durasi kedua jenis ekspresi main-main dinilai menggunakan rata-rata durasi per kejadian kesepakatan antar pengamat (IOA) untuk sejumlah besar waktu (Reed dan Azulay 2010 ). Kami memilih sekitar 10% PF (durasi 40 PF yang dinilai) dan FPF (durasi 90 FPF yang dinilai) yang direkam pada video untuk menghitung IOA. Durasi setiap PF/FPF (yaitu, uji coba pengukuran durasi) dievaluasi oleh kedua pengamat secara terpisah. IOA ditentukan untuk setiap uji coba pengukuran durasi dengan membagi durasi yang lebih kecil dengan durasi yang lebih tinggi yang dilaporkan oleh kedua pengamat. Misalnya, jika durasi PF adalah 15 detik untuk satu pengamat dan 20 detik untuk pengamat lainnya, IOA untuk uji coba pengukuran durasi ini adalah 15/20 = 0,75. Nilai IOA dari semua uji coba pengukuran durasi dijumlahkan dan dibagi dengan jumlah total uji coba; hasilnya diubah dalam %. IOA untuk durasi PF dan FPF sama dengan 93%.
2.5.3 Evaluasi Keberadaan RFM
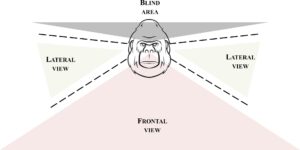
Untuk menunjukkan keberadaan RFM dalam kelompok yang diteliti, setiap kali seorang pemain (selanjutnya disebut pemicu) memancarkan PF/FPF pertama (selanjutnya disebut stimulus pertama), kami mengevaluasi keberadaan atau ketiadaan PF/FPF yang dipancarkan oleh partner bermain (selanjutnya disebut responder potensial) dalam 1 detik setelah emisi stimulus pertama di bawah dua kondisi persepsi (Gambar 2 ): stimulus pertama dipersepsikan oleh responder potensial (yaitu, stimulus jatuh dalam medan visual responder; kondisi persepsi ya ) dan stimulus pertama tidak dipersepsikan oleh responder potensial (yaitu, stimulus tidak jatuh dalam medan visual responder; kondisi tidak ada persepsi ). Semua kasus yang meragukan (misalnya, ketika pengamat tidak melihat dengan jelas wajah pemain dari video, atau posisi kepala pemain tidak jelas) dibuang dari analisis. Kesepakatan antara kedua pengamat dalam menentukan kondisi persepsi ya dan tidak persepsi diukur dalam 300 sesi permainan (sekitar 30% dari total sesi permainan yang dianalisis), termasuk 15 gorila untuk persepsi ya-/tidak terhadap PF dan 12 gorila untuk persepsi ya-/tidak terhadap FPF. Kappa Cohen mencapai nilai 0,82 untuk persepsi ya-/tidak terhadap PF dan 0,83 untuk persepsi ya-/tidak terhadap FPF. Kami melengkapi analisis ini dengan analisis lain yang mempertimbangkan berapa banyak PF/FPF yang dipancarkan oleh pemicu yang dipersepsikan vs tidak dipersepsikan oleh responden potensial dan berapa banyak dari PF/FPF ini yang direplikasi atau tidak direplikasi oleh responden potensial dalam 1 detik setelah emisinya. Berdasarkan definisi RFM (lihat Bagian 1 ), semua analisis ini hanya mencakup respons kongruen oleh responden potensial terhadap stimulus pertama: PF sebagai respons terhadap PF dan FPF sebagai respons terhadap FPF (pencocokan wajah yang tepat).
Kami kemudian membandingkan tingkat individu PF_RFM dengan tingkat individu FPF_RFM dengan membagi jumlah PF/FPF yang direplikasi oleh responden potensial dengan jumlah PF/FPF yang mereka rasakan.
2.5.4 Durasi Sesi Bermain
Bahasa Indonesia: Untuk mengevaluasi kemungkinan efek PF, FPF, atau RFM pada durasi sesi permainan, kami mempertimbangkan lima kondisi berbeda: (i) pemain melakukan setidaknya dua PF yang tidak ditiru saja ( PF-not-mim ); (ii) pemain melakukan setidaknya dua FPF yang tidak ditiru saja ( FPF-not-mim ); (iii) setidaknya satu kejadian RFM PF (tetapi bukan FPF) terjadi ( PF_RFM ); (iv) setidaknya satu kejadian RFM FPF (tetapi bukan PF) terjadi ( FPF_RFM ), dan (v) setidaknya satu kejadian RFM PF dan RFM FPF terjadi dalam sesi permainan yang sama ( PF&FPF_RFM ). Untuk setiap kondisi, kami menentukan periode waktu (dalam detik) antara terjadinya kejadian PF, FPF, atau RFM pertama dan akhir sesi permainan (yaitu, periode waktu yang tersisa dari PF/FPF/RFM pertama). Kemudian, kami membandingkan periode waktu ini di kelima kondisi. Kami mengecualikan dari analisis tersebut sesi bermain di mana tidak ada peristiwa PF/FPF atau RFM terjadi, serta kasus ambigu di mana PF dan FPF yang tidak ditiru keduanya hadir dalam sesi yang sama. Lebih jauh, untuk menentukan apakah RFM dari PF atau FPF terjadi di dekat awal (untuk memperpanjang sesi bermain) atau akhir (untuk menandakan akhir sesi bermain) dari sesi bermain, kami menghitung titik tengah durasi (dalam detik) dari setiap sesi. Kami kemudian mengevaluasi apakah peristiwa RFM pertama terjadi sebelum (mendekati awal sesi) atau setelah (mendekati akhir sesi) titik tengah ini (variabel binomial: sebelum titik tengah = 1; setelah titik tengah = 0).
2.5.5 Indeks Asimetri Permainan (PAI)
PAI dihitung dengan mempertimbangkan jumlah pola perilaku menyerang dan bertahan (lihat Tabel 2 ) yang dipertukarkan antara pemain pada jumlah total pola (menyerang + bertahan + netral) yang menyusun sesi (Cordoni et al. 2018 , 2023 ):
![]()
Dalam rumus, A dan B mewakili pemain dan panah (
) yang mengarahkan perilaku kepada siapa. PAI dihitung sebelum dan setelah peristiwa PF/FPF atau RFM (lihat Bagian 2.5.7 ) dan berkisar dari -1 hingga +1. Nilai PAI yang sama dengan atau mendekati -1 menunjukkan asimetri lengkap yang menguntungkan Pemain B, yang melakukan lebih banyak pola ofensif/lebih sedikit pola defensif atau menerima lebih sedikit pola ofensif/lebih banyak pola defensif dari Pemain A. Nilai PAI yang sama dengan atau mendekati +1 menunjukkan asimetri lengkap yang menguntungkan Pemain A. Nilai PAI yang sama dengan atau mendekati 0 menunjukkan simetri lengkap dalam pertukaran perilaku ofensif dan defensif antara pemain.
2.5.6 Indeks Shannon ( H ′)
H ′ (Shannon 1948 ; Keylock 2005 ) adalah indeks ekologi yang diadaptasi untuk mengevaluasi tingkat variabilitas permainan dalam hal berbagai jenis pola perilaku yang dilakukan oleh pemain selama sesi (Cordoni et al. 2023 ). Semua pola ofensif, defensif, dan netral yang dijelaskan dalam Tabel 2 dipertimbangkan untuk perhitungan H ′. Indeks dihitung sebagai berikut:
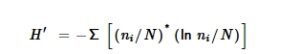
Dalam rumus, n i mewakili jumlah perilaku yang termasuk dalam tipe i dan N mewakili jumlah total perilaku yang menyusun sesi. Misalnya, untuk sesi yang disusun oleh “bermain menampar—bermain menampar—bermain berlindung—bermain menampar—berputar,” n i = 3 (tipe perilaku = bermain menampar , bermain berlindung , dan berputar ), N = 5 (jumlah total pola perilaku yang menyusun sesi yaitu 3 bermain menampar + 1 bermain berlindung + 1 berputar ). Nilai H ′ yang tinggi menunjukkan variabilitas pola perilaku yang besar.
2.5.7 Analisis Sekuensial
Bahasa Indonesia: Untuk mengevaluasi kemungkinan variasi dalam asimetri permainan dan variabilitas dalam sesi yang terkait dengan kemunculan PF, FPF, atau RFM, kami melakukan analisis berurutan pada setiap sesi permainan, seperti yang dirinci dalam lembar Excel (lihat Pengumpulan Data). Ketika peristiwa PF, FPF, atau RFM (baik PF maupun FPF) terjadi dalam sesi, kami memilih—jika memungkinkan—empat pola permainan sebelum dan empat pola permainan setelah ekspresi main-main (PF atau FPF) atau peristiwa mimikri (PF_RFM atau FPF_RFM). Berdasarkan pola-pola ini, kami menghitung nilai PAI dan H ′ sebelum dan sesudah peristiwa PF, FPF, atau RFM. Akibatnya, untuk setiap peristiwa PF, FPF, atau RFM dalam sesi, kami memperoleh PAI sebelum , PAI sesudah , H ′ sebelum , dan H ′ sesudah . Kami memilih empat pola sebelum dan sesudah ekspresi main-main atau peristiwa RFM untuk menstandardisasi data dan memberikan probabilitas kepada setiap pemain untuk melakukan setidaknya satu pola ofensif dan satu pola defensif. Lebih jauh, karena mungkin ada bias durasi sesi (variabilitas tinggi dalam durasi sesi), kami membakukan analisis ini dengan memilih empat pola sebelum dan empat pola sesudahnya. Ketika beberapa ekspresi main-main atau peristiwa RFM terjadi secara berurutan dalam satu sesi, kami menghitung PAI sebelum , PAI sesudah , H ′ sebelum , dan H ′ sesudah hanya jika dua ekspresi atau peristiwa RFM dipisahkan oleh setidaknya empat pola main (tidak termasuk empat pola yang digunakan untuk perhitungan indeks). Jadi, kami mengecualikan dari analisis ekspresi main-main atau peristiwa RFM yang berurutan yang dipisahkan oleh kurang dari empat pola.
2.6 Analisis Statistik
Distribusi periode waktu dalam lima kondisi (lihat Bagian 2.5 ) tidak normal (uji Shapiro–Wilk 0,643 ≤ W ≤ 0,862; 0,001 ≤ p ≤ 0,029); dengan demikian, kami menggunakan uji Kruskal–Wallis non-parametrik yang digunakan untuk sampel k -independen. Kami juga menerapkan pengacakan Monte Carlo (10.000 permutasi) karena data tidak independen karena fakta bahwa individu yang sama dapat hadir dalam lebih dari satu dyad. Dalam kasus signifikansi uji, kami menerapkan uji post hoc dengan koreksi Bonferroni untuk perbandingan berpasangan.
Durasi median PF/FPF, nilai intensitas AU, kadar PF_RFM dan FPF_RFM, dan nilai PAI/ H ′ (uji Shapiro–Wilk 0,612 ≤ W ≤ 0,800; 0,001 ≤ p ≤ 0,007) dibandingkan pada tingkat individu dengan menggunakan uji eksak Wilcoxon non-parametrik untuk dua sampel dependen dengan koreksi Bonferroni jika diperlukan.
Kami menggunakan uji binomial untuk mengevaluasi apakah kejadian RFM pertama dalam sesi terjadi lebih sering sebelum atau setelah titik tengah durasi (dalam detik) setiap sesi.
Uji chi-square kontingensi 2 × 2 digunakan untuk membandingkan jumlah PF/FPF yang direplikasi atau tidak direplikasi dalam 1 detik oleh responden potensial dalam kondisi yang dipersepsikan dan tidak dipersepsikan. Untuk memastikan hasil yang lebih konservatif, Koreksi Kontinuitas Yates diterapkan.
Untuk membandingkan jumlah individu PF/FPF yang dilakukan per sesi bermain (uji Shapiro–Wilk 0,146 ≤ W ≤ 0,183; 0,066 ≤ p ≤ 0,200), kami menggunakan uji- t berpasangan parametrik untuk sampel dependen.
Untuk menunjukkan kemunculan RFM untuk PF dan FPF, kami menjalankan dua model campuran linear tergeneralisasi (GLMM). Model pertama (GLMM PF ) mencakup sebagai variabel target kehadiran/ketiadaan (variabel binomial; 0 = ketidakhadiran, 1 = kehadiran) PF yang dilakukan oleh responden potensial dalam 1 detik setelah emisi PF pertama oleh pemicu. Faktor tetap adalah kondisi persepsi responden potensial (variabel binomial; 0 = tidak ada persepsi , 1 = ya-persepsi ) dan kelompok (variabel binomial; 1 = kelompok BEA, 2 = kelompok VDS). ID dyad (pemicu-responden potensial) dimasukkan sebagai faktor acak.
Model kedua (GLMM FPF ) memasukkan sebagai variabel target ada/tidaknya (variabel binomial; 0 = tidak ada, 1 = ada) FPF yang dilakukan oleh responden potensial dalam 1 detik setelah emisi FPF pertama oleh pemicu. Faktor tetap adalah kondisi persepsi responden potensial (variabel binomial; 0 = tidak ada persepsi , 1 = ya-persepsi ) dan kelompok (variabel binomial; 1 = kelompok BEA, 2 = kelompok VDS). ID dyad (pemicu-responden potensial) dimasukkan sebagai faktor acak.
3 Hasil
3.1 Analisis Awal
Kami melakukan analisis ini pada tingkat individu dengan membandingkan jumlah PF/FPF yang dilakukan per sesi permainan (yaitu, jumlah total PF/FPF yang dilakukan oleh seorang individu dibagi dengan jumlah total sesi permainan yang melibatkan individu tersebut). Kami menemukan bahwa kadar FPF lebih tinggi daripada kadar PF (paired t -test N individu = 21, gl = 20, t = −2.608, p = 0,017; nilai rata-rata PF/FPF per sesi ± SE: PF 0,26 ± 0,05; FPF 0,51 ± 0,11; Gambar 3A ).

Untuk setiap individu, kami juga menghitung median durasi PF dan FPF. Durasi median tidak berbeda secara signifikan antara PF dan FPF (uji eksak Wilcoxon N individu = 16, T = 56,0, seri = 1, p = 0,835; Gambar 3B ). Ukuran sampel adalah 16 karena analisis ini hanya mencakup individu yang PF dan FPF-nya tercatat (lihat Tabel 1 ).
Persentase sesi bermain di mana hanya satu jenis ekspresi main-main dilakukan masing-masing adalah 3,1% dan 5,5% untuk PF dan FPF.
3.2 Prediksi 1
Melalui penggunaan chimpFACS, selama pelaksanaan PF, kami mendeteksi aktivasi AU yang dilaporkan dalam Tabel S2 dan Gambar S3 . Kami memperoleh 14 konfigurasi berbeda untuk PF dan—meskipun ukuran sampel terbatas ( N = 8 gorila)—kami menemukan bahwa empat AU selalu atau hampir selalu hadir dalam konfigurasi yang terdeteksi: AU12 ( zygomaticus mayor —penarik sudut bibir), AU16 ( depressor labii inferioris —depresor bibir bawah), AU25 ( depressor labii , orbicularis oris —bagian bibir), dan AU26 (otot non-mimetik—rahang jatuh). Kami juga melaporkan dalam Tabel S2 dan Gambar S4 aktivasi AU selama pelaksanaan FPF ( N = 9 gorila). Kami memperoleh enam konfigurasi berbeda untuk FPF dan delapan AU selalu atau hampir selalu hadir dalam konfigurasi yang terdeteksi: AU06 ( orbicularis oculi , pars orbitalis —pengangkat pipi), AU09 ( levator labii superioris alaquae nasi —pengerut hidung), AU10 ( levator labii superioris —pengangkat bibir atas), AU12, AU16, AU25, AU26, dan AU27 (otot non-mimetik—peregangan mulut). AU10 diaktifkan hanya dua kali dan AU27 hanya sekali selama pelaksanaan PF, sehingga menunjukkan bahwa PF dan FPF mungkin berbeda sampai batas tertentu dalam pengangkatan bibir atas dan peregangan mulut.
Dengan OpenFace 2.0, kami memperoleh nilai intensitas AU yang terlibat dalam kinerja PF dan FPF (lihat data mentah yang disediakan sebagai Informasi Pendukung ). OpenFace tidak memperkirakan nilai intensitas AU16 dan AU27; karena alasan ini, analisis berikut tidak menyertakan kedua AU ini. Bahasa Indonesia:Untuk setiap AU, kami membandingkan nilai intensitas antara PF dan FPF pada tingkat individu, dan kami memperoleh hasil berikut (koreksi Bonferroni α = 0,005): AU1—pengangkat alis bagian dalam (uji pasti Wilcoxon N individu = 8, T = 4, ikatan = 2, p = 0,219), AU2 pengangkat alis bagian luar ( N individu = 8, T = 0, ikatan = 0, p = 1,000), AU6 ( N individu = 8, T = 10,5, ikatan = 0, p = 0,328), AU9 ( N individu = 8, T = 5, ikatan = 3, p = 0,498), AU10 ( N individu = 8, T = 0, ikatan = 0, p = 0,008), AU12 ( N individu = 8, T = 4,5, ikatan = 0, p = 0,063), AU17—chin raiser ( N individu = 8, T = 3, seri = 0, p = 0,039), AU 25 ( N individu = 8, T = 10, seri = 0, p = 0,313), AU 26 ( N individu = 8, T = 11, seri = 0, p = 0,383), dan AU45—blink ( N individu = 8, T = 13,5, seri = 1, p = 0,984). Meskipun tidak ada perbandingan yang mencapai signifikansi statistik, probabilitas AU10 adalah yang paling dekat dengan signifikansi ketika mempertimbangkan koreksi Bonferroni. Kita dapat menyimpulkan bahwa intensitas AU10 cenderung lebih tinggi selama FPF daripada kinerja PF.
Oleh karena itu, hasil dari chimpFACS dan OpenFace manusia menunjukkan bahwa PF dan FPF mungkin berbeda terutama dalam aktivasi AU10 (mengangkat bibir lebih tinggi) dan AU27 (peregangan mulut) yang terutama bertanggung jawab atas tereksposnya gigi atas.
3.3 Prediksi 2
Prediksi 2a . Kami menjalankan GLMM PF untuk memverifikasi kemunculan PF_RFM. Model lengkap (termasuk faktor tetap) dan model nol (hanya termasuk faktor acak) berbeda secara signifikan (uji rasio kemungkinan: χ 2 = 8,281, df = 2, p = 0,016). Karena prediktor memiliki efek signifikan pada variabel target, kami menerapkan prosedur drop1. Kami menemukan bahwa kondisi persepsi secara signifikan memengaruhi variabel target: probabilitas emisi PF oleh responden potensial lebih tinggi ketika mereka secara visual mempersepsikan PF yang dipancarkan oleh pemicu ( kondisi persepsi ya ) dibandingkan dengan ketika mereka tidak secara visual mempersepsikan ( kondisi tidak ada persepsi ) stimulus pertama (Tabel 3 ). Uji chi-kuadrat kontingensi 2 × 2 menunjukkan bahwa PF secara signifikan lebih banyak direplikasi ketika dirasakan oleh responden potensial dibandingkan dengan kondisi lainnya ( χ 2 = 5.080, df = 1, p = 0,024; Gambar 4A ).
| Memperkirakan | Bahasa Inggris | nilai z | P | |
|---|---|---|---|---|
| Variabel target PF GLMM : ada/tidaknya PF oleh responden potensial dalam 1 detik setelah stimulus pertama oleh pemicu (hanya respons kongruen yang dianggap PF→PF)
Faktor acak: ID dyad ( N dyad = 117) model penuh versus model nol: χ 2 = 8.281, gl = 2, p = 0.016 |
||||
| Mencegat | -2,373 | 0.854 | -2.780 | 0,005 |
| Kondisi persepsi ( ya-persepsi ) | 1.064 | 0.463 | 2.296 | 0,022 |
| Grup (VDS) | 1.109 | 0.814 | 1.363 | 0.173 |
| Faktor Inflasi Varians (VIF) | ||||
| Kondisi persepsi | 1.001 | |||
| Kelompok | 1.001 | |||
| Variabel target FPF GLMM : ada/tidaknya FPF oleh responden potensial dalam 1 detik setelah stimulus pertama oleh pemicu (hanya respons kongruen yang dianggap FPF→FPF)
Faktor acak: ID dyad ( N dyad = 343) model penuh versus model nol: χ 2 = 50.239, gl = 2, p < 0,001 |
||||
| Mencegat | -1.469 | 0,562 | -2.615 | 0,009 |
| Kondisi persepsi ( ya-persepsi) ) | 2.117 | 0,338 | 6.259 | < 0,001 |
| Grup (VDS) | -0,157 | 0,501 tahun | -0,313 | 0,755 tahun |
| Faktor Inflasi Varians (VIF) | ||||
| Kondisi persepsi | 1.003 | |||
| Kelompok | 1.003 | |||
Catatan: Faktor acak = pasangan yang terdiri dari pemicu dan calon responden. Nilai yang dicetak tebal menunjukkan signifikansi statistik ( p < 0,05). Prediktor diberi kode dummy, dengan kategori referensi kondisi persepsi ( ya-persepsi) ) dan kelompok (VDS). Singkatan: SE, kesalahan standar; VDS, La Vallée des Singes.

GLMM FPF dilakukan untuk memverifikasi kemunculan FPF_RFM. Model penuh dan model nol berbeda secara signifikan (uji rasio kemungkinan: χ 2 = 50,239, df = 2, p < 0,001). Kami menemukan bahwa kondisi persepsi secara signifikan memengaruhi variabel target: probabilitas emisi FPF oleh responden potensial lebih tinggi ketika mereka secara visual mempersepsikan FPF yang dipancarkan oleh pemicu ( kondisi persepsi ya ) dibandingkan ketika mereka tidak mempersepsikan secara visual ( kondisi tidak ada persepsi ) stimulus pertama (Tabel 3 ). Uji chi-kuadrat kontingensi 2 × 2 menunjukkan bahwa FPF secara signifikan lebih banyak direplikasi ketika dipersepsikan oleh responden potensial dibandingkan dengan kondisi lainnya ( χ 2 = 45,024, df = 1, p < 0,001; Gambar 4B ). Singkatnya, RFM ditentukan untuk PF dan FPF secara terpisah.
Tingkat FPF_RFM cenderung lebih tinggi daripada tingkat PF_RFM (uji pasti Wilcoxon N = 10, T = 7,0, ikatan = 1,0, p = 0,074; tingkat rata-rata RFM ± SE: PF_RFM 0,31 ± 0,08; FPF_RFM 0,57 ± 0,10). Ukuran sampel lebih kecil ( N = 10) karena analisis hanya mencakup individu yang telah merasakan dan meniru PF dan FPF.
Prediksi 2b . Periode waktu (dalam detik) antara kemunculan ekspresi main-main pertama (PF/FPF) atau peristiwa RFM dan akhir sesi bermain berbeda secara signifikan di lima kondisi yang dipertimbangkan (uji Kruskal–Wallis—pengacakan Monte Carlo N periode waktu = 186, H = 21,711, df = 4, p < 0,001; Gambar 5 ). Uji post hoc menunjukkan hasil berikut: PF-bukan-mim versus FPF-bukan-mim (uji post hoc Bonferroni—Dunn Q = −4,953, p = 1,000), PF-bukan-mim versus PF_RFM ( Q = −21,466, p = 1,000), PF-bukan-mim versus FPF_RFM ( Q = −35,570, p = 0,021), PF-bukan-mim versus PF&FPF_RFM ( Q = −63,891, p = 0,007), FPF-bukan-mim versus PF_RFM ( Q = −16,513, p = 1,000), FPF-bukan-mim versus FPF_RFM ( Q = −30,617, p = 0,017), FPF-bukan-mim versus PF&FPF_RFM ( Q = −58,938, p = 0,009), PF_RFM versus FPF_RFM ( Q = −14,104, p = 1,000), PF_RFM versus PF&FPF_RFM ( Q = −42,425, p = 0,358), dan FPF_RFM versus PF&FPF_RFM ( Q = −28,231, p = 1,000). Dengan demikian, periode waktu dari terjadinya peristiwa RFM pertama yang melibatkan FPF atau PF-FPF hingga akhir sesi permainan lebih panjang dibandingkan dengan periode waktu setelah PF atau FPF pertama yang tidak ditiru.
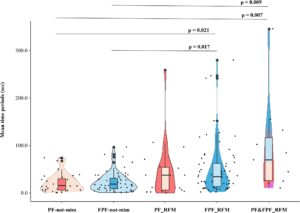
Kami juga menemukan bahwa peristiwa RFM baik PF maupun FPF terjadi lebih sering sebelum daripada setelah titik tengah durasi (dalam detik) setiap sesi (Uji Binomial N RFMevents = 187, p < 0,001).
3.4 Prediksi 3
Prediksi 3a . Nilai PAI tidak berbeda secara signifikan sebelum dan setelah emisi PF-bukan-mim (uji Wilcoxon—pengacakan Monte Carlo N kejadian = 30, T = 53, ikatan = 15, p = 0,685; nilai PAI rata-rata ± SE: sebelum 0,333 ± 0,060; setelah 0,308 ± 0,050), kejadian PF_RFM ( N kejadian = 11, T = 11,5, ikatan = 3, p = 0,449; sebelum 0,409 ± 0,127; setelah 0,273 ± 0,071) dan kejadian FPF_RFM ( N kejadian = 29, T = 88,5, ikatan = 10, p = 0,780; sebelum 0,362 ± 0,060; setelah 0,362 ± 0,055). Sebaliknya, kami menemukan bahwa nilai PAI lebih tinggi setelah daripada sebelum emisi FPF -bukan-mim ( N kejadian = 86, T = 847, ikatan = 18, p = 0,037; sebelum 0,352 ± 0,037; setelah 0,474 ± 0,038; Gambar 6A ). Oleh karena itu, hanya kemunculan FPF yang tidak ditiru yang diikuti oleh peningkatan asimetri pertukaran pola permainan yang berurutan.

Kami membandingkan semua nilai PAI sebelum (uji Kruskall–Wallis, pengacakan Monte Carlo N events_PF_not_mimicked = 30, N events_FPF_not_mimicked = 86, N events_PF_RFM = 11, N events_FPF_RFM = 29, H = 0,277, df = 3, p = 0,964) dan sesudah (uji Kruskall–Wallis, pengacakan Monte Carlo N events_PF_not_mimicked = 30, N events_FPF_not_mimicked = 86, N events_PF_RFM = 11, N events_FPF_RFM = 29, H = 7,497, df = 3, p = 0,060) PF-not-mim , FPF-not-mim , PF_RFM , dan FPF_RFM , dan kami tidak memperoleh perbedaan statistik apa pun dalam kedua kasus.
Prediksi 3b . Kami tidak menemukan perbedaan signifikan dalam nilai Indeks Shannon ( H ′) sebelum dan setelah emisi FPF -bukan-mim (uji Wilcoxon—pengacakan Monte Carlo N kejadian = 86, T = 918,5, ikatan = 25, p = 0,851; nilai H ′ rata-rata ± SE: sebelum 0,600 ± 0,054; setelah 0,610 ± 0,053), kejadian PF_RFM ( N kejadian = 11, T = 10,5, ikatan = 3, p = 0,345; sebelum 0,626 ± 0,150; setelah 0,400 ± 0,105) dan kejadian FPF_RFM ( N kejadian = 29, T = 92,5, ikatan = 10, p = 0,937; sebelum 0,663 ± 0,100; setelah 0,700 ± 0,100). Sebaliknya, nilai H ′ secara signifikan lebih tinggi setelah daripada sebelum emisi PF-bukan-mim ( N peristiwa = 30, T = 49,0, ikatan = 10, p = 0,034; sebelum 0,530 ± 0,100; setelah 0,815 ± 0,100; Gambar 6B ). Oleh karena itu, hanya kemunculan PF yang tidak ditiru yang diikuti oleh peningkatan variabilitas pola permainan yang berurutan.
Kami membandingkan semua nilai H ′ sebelum (uji Kruskall–Wallis, pengacakan Monte Carlo N events_PF_not_mimicked = 30, N events_FPF_not_mimicked = 86, N events_PF_RFM = 11, N events_FPF_RFM = 29, H = 3,201, df = 3, p = 0,367) dan sesudah (uji Kruskall–Wallis, pengacakan Monte Carlo N events_PF_not_mimicked = 30, N events_FPF_not_mimicked = 86, N events_PF_RFM = 11, N events_FPF_RFM = 29, H = 6,902, df = 3, p = 0,072) PF-not-mim , FPF-not-mim , PF_RFM , dan FPF_RFM , dan kami tidak memperoleh perbedaan statistik apa pun dalam kedua kasus.
Analisis pada PAI dan H ′ mencakup 100 sesi bermain.
4 Diskusi
Menurut Hipotesis Kompleksitas dan Kontinuitas (Davila-Ross dan Dezecache 2021 ), pada manusia, tawa dan senyuman yang menunjukkan afek positif pasti telah berevolusi dalam konteks bermain pada spesies leluhur, dan ekspresi ini telah melalui periode utama perubahan evolusi melalui jalur filogenetik yang berbeda dalam 10–16 juta tahun terakhir hingga menjadi pola yang efektif, luas, dan fleksibel yang digunakan selama interaksi sosial pada Hominidae (Davila-Ross dan Zimmermann 2009 ; Davila-Ross et al. 2008 , 2015 ; Waller et al. 2015). ).
Waller et al. ( 2020 ) berpendapat bahwa untuk mempertimbangkan ekspresi wajah sebagai homologus di seluruh spesies (termasuk manusia) diperlukan untuk menunjukkan “bentuk yang stereotip dan dapat dikenali…, kesamaan beberapa elemen, homologi otot wajah yang mendasarinya dan kehadiran dalam sejumlah besar spesies terkait” (9).
Dalam studi terkini, kami menunjukkan bahwa pada gorila dataran rendah, PF dan FPF tampaknya mempertahankan kesamaan morfologi (“homologi otot wajah yang mendasarinya”) dengan PF dan FPF yang ditampilkan oleh kera besar lainnya (“kehadiran dalam sejumlah besar spesies terkait”; Prediksi 1a didukung; lihat Video S1A,B dan Tabel S2 ) sampai pada titik di mana alat yang dirancang untuk mendeteksi aktivasi AU pada manusia dan simpanse juga bekerja dengan kesalahan minimal untuk gorila (GorillaFACS baru-baru ini diimplementasikan oleh Correia-Caeiro et al. 2025 , dan belum tersedia pada saat studi kami).
Dalam penelitian kami terhadap gorila, kinerja PF melibatkan sebagian besar AU (Tabel S2 ) yang dijelaskan untuk PF simpanse, di mana dua konfigurasi terbaik mencakup AU12 + AU25 + AU26 dan AU12 + AU25 + AU27, dengan AU26 dan AU27 saling eksklusif (Parr et al. 2007 ). Selain itu, pada orangutan, AU26 dan AU27 terlibat dalam kinerja PF, dengan AU27 lebih aktif daripada AU26 selama sesi bermain dengan intensitas lebih tinggi (Waller et al. 2015 ). Di sisi lain, pada gorila dataran rendah, kinerja FPF—lebih dari PF—melibatkan aktivasi AU (Tabel S2 ) yang bertanggung jawab untuk mengangkat bibir atas (AU10) dan meregangkan mulut (AU27), mirip dengan ekspresi wajah lebar dan ceria yang diamati pada simpanse (Vick et al. 2007 ). Pada manusia, wajah tertawa dan senyum (kemungkinan homolog PF dan FPF) mencakup gerakan otot wajah yang mirip dengan kera besar lainnya, seperti aktivasi zygomaticus mayor (AU12) dan orbicularis oculi (AU06) yang menyebabkan pipi terangkat dan mata berkerut (Ruch dan Ekman 2001 ; Drack et al. 2009 ; Masai et al. 2022). ).
Temuan kami menunjukkan bahwa pada gorila, PF dan FPF tidak sepenuhnya tumpang tindih dari sudut pandang morfologi ( Prediksi 1b didukung; lihat Tabel S2 ). Seperti yang diharapkan, sebaliknya terhadap PF, unit otot yang terkait dengan paparan gigi yang lebih tinggi selalu terlibat dalam kinerja FPF. Meskipun terjadi pada frekuensi rendah, PF dan FPF dapat hadir secara independen satu sama lain selama interaksi bermain (3,1% dan 5,5% dari sesi bermain dengan hanya PF dan FPF yang dilakukan, masing-masing) dan, untuk pertama kalinya, kami menunjukkan bahwa PF atau FPF dapat dengan cepat direplikasi oleh pemain (RFM; Prediksi 2a didukung; Gambar 4 ). Meskipun tingkat FPF_RFM cenderung lebih tinggi daripada PF_RFM, signifikansi statistik tidak tercapai. Data lebih lanjut diperlukan untuk menilai secara definitif setiap perbedaan potensial dalam tingkat replikasi antara PF dan FPF. Sejauh pengetahuan kami, keberadaan RFM pada primata nonmanusia sejauh ini ditemukan dengan menggabungkan PF dan FPF (misalnya, orangutan, Davila-Ross et al. 2008 ; gorila dataran rendah dan simpanse, Palagi, Norscia, et al. 2019 ; Bresciani et al. 2021 ; bonobo, Bertini et al. 2021 ). Pada gorila, RFM dari PF dan FPF dapat berfungsi dalam membina ikatan sosial antara individu selama konteks bermain yang positif (dan aman) (Bresciani et al. 2021 ). Juga pada manusia, hanya senyum dan tawa yang dianggap sebagai sinyal positif dan tepat yang dapat meningkatkan afiliasi antara individu dan peningkatan ini dapat diekspresikan melalui mimikri wajah (Kastendieck et al. 2021 ). Berbagi tawa mengomunikasikan pemahaman bersama tentang konteks tersebut (Martin dan Ford 2018 ).
Pada gorila kami, kami menemukan bahwa periode waktu (diukur dalam detik) dari terjadinya peristiwa RFM pertama yang melibatkan FPF atau PF-FPF hingga akhir sesi bermain lebih lama dibandingkan dengan periode waktu setelah PF atau FPF pertama yang tidak ditiru ( Prediksi 2b sebagian didukung; Gambar 5 ). Dengan demikian, adanya replikasi wajah, bukan hanya ekspresi wajah yang tidak ditiru, dapat berkontribusi untuk memperpanjang sesi bermain. Selain itu, sebagian besar peristiwa RFM terjadi di dekat awal sesi, yang menunjukkan bahwa peristiwa tersebut tidak menandai akhir dari interaksi bermain. RFM dapat meningkatkan sinkronisasi tindakan dan berbagi suasana hati bermain antara pemain (terutama pemain yang ukurannya cocok; Bresciani et al. 2021 ) sehingga mendukung pemeliharaan interaksi. Pada manusia, interaksi sosial yang berkepanjangan dapat dikaitkan dengan penularan tawa yang bertindak sebagai perekat sosial (Provine 1992 ; Dunbar 2022 ). Namun, dalam kelompok gorila kami, kami menemukan bahwa PF_RFM dapat memperpanjang sesi bermain terutama jika terjadi bersamaan dengan FPF_RFM, sehingga menghasilkan efek yang lebih kuat. Kami dapat berhipotesis bahwa replikasi sinyal yang lebih kentara, seperti FPF, dapat mengomunikasikan maksud untuk melanjutkan permainan dengan lebih jelas, sehingga meminimalkan kemungkinan salah tafsir dan mencegah permainan meningkat menjadi konflik (Palagi et al. 2007 ; Waller dan Cherry 2012 ).
Kami menunjukkan bahwa FPF—tetapi bukan PF atau RFM dari PF atau FPF—segera diikuti oleh peningkatan tingkat asimetri permainan dalam sesi ( Prediksi 3a didukung; Gambar 4A ). Semua sesi bermain yang kami analisis melibatkan kontak fisik antara pemain (sesi perkelahian bermain) dan tingkat FPF per sesi lebih tinggi dibandingkan dengan PF (Gambar 3A ), bahkan jika kedua sinyal dipertahankan untuk jangka waktu yang sebanding (Gambar 3B ). Penggunaan sinyal yang lebih jelas (FPF) yang lebih sering dapat mengomunikasikan maksud baik dari agen dengan lebih baik. Akibatnya, pernyataan tujuan yang jelas dapat memungkinkan subjek untuk mengubah permainan menjadi interaksi yang lebih kompetitif dan menuntut kognitif, yang dapat meningkatkan fungsi penilaian diri dan sosial dari perilaku bermain (Fagen 1981 ; Paquette 1994 ; Thompson 1998 ). Menurut Teori Polyvagal , bermain dapat dilihat sebagai latihan saraf karena memerlukan kapasitas untuk beralih antara respons kompetitif melawan/lari dan interaksi sosial kooperatif (Porges 2011 ). Dalam hal ini, FPF dapat mendorong transisi antara kerja sama/kompetisi dalam sesi dengan cara yang cepat dengan mempertahankan konteks bermain yang “tidak serius”. Demikian pula, pada anak-anak prasekolah, tawa yang berlebihan (sinyal yang lebih jelas) paling sering dikaitkan dengan agresi pura-pura, yang merupakan bentuk interaksi bermain yang berisiko (Sarra dan Otta 2001 ). Dengan demikian, dari sudut pandang evolusi, kita dapat berasumsi bahwa penggunaan sinyal yang lebih jelas dipertahankan ketika diperlukan untuk memperoleh respons perilaku yang benar oleh pasangan dan ketika berbagi konteks (misalnya, interaksi bermain intensitas tinggi) mungkin penting untuk membatasi risiko kesalahpahaman (“kesamaan beberapa elemen”).
Berdasarkan Hipotesis Otak Sosial (Dunbar 2022 ; Shultz dan Dunbar 2012 ) komunikasi berfungsi sebagai perekat sosial antara anggota kelompok. FPF pada gorila dataran rendah mungkin memiliki efek sosial yang positif dengan meningkatkan penilaian sosial pemain, seperti menciptakan dan meningkatkan ikatan antara individu (Bresciani et al. 2021 ). Baru-baru ini telah ditunjukkan bahwa pada manusia, orang yang lebih ekspresif pada wajah lebih menyenangkan dan disukai oleh pasangan sosialnya, sehingga menunjukkan bahwa ekspresi wajah dapat mendukung pembentukan dan pemeliharaan hubungan sosial antar individu (Kavanagh et al. 2022 ). Kita dapat berasumsi bahwa efek sosial dari komunikasi wajah ini adalah hasil dari kesinambungan evolusi dari hominid non-manusia hingga manusia modern.
Kami juga menunjukkan bahwa PF—tetapi bukan FPF dan RFM dari PF atau FPF—diikuti oleh peningkatan variabilitas sesi bermain ( Prediksi 3b mendukung). Dalam pandangan ini, PF dapat mendorong penggunaan pola bermain yang lebih berbeda dalam jangka pendek oleh pemain yang menghasilkan sesi bermain yang lebih bervariasi—dan akibatnya lebih menuntut secara fisik dan kognitif. Meningkatnya penggunaan berbagai jenis pola bermain dalam satu sesi dapat memfasilitasi pelatihan untuk hal yang tidak terduga (Spinka et al. 2001 ), karena pemain tidak dapat mengantisipasi perilaku pasangannya tetapi harus mampu merespons pola bermain yang tidak terduga dengan tepat.
4.1 Batasan Generalitas (Simons et al. 2017 ) dan Kesimpulan
Meskipun penelitian kami hanya berfokus pada 21 gorila dataran rendah dalam penangkaran dan kehati-hatian diperlukan saat menggeneralisasi temuan ini, ini mungkin upaya pertama untuk menganalisis PF dan FPF dan replikasi cepatnya secara terpisah. Kami yakin bahwa hasil kami dapat mewakili titik awal untuk mendorong lebih banyak penelitian tentang perbedaan potensial antara dua varian ekspresi wajah yang menyenangkan. Meskipun hanya 17 PF dan 15 FPF yang dinilai dengan menggunakan alat chimpFACS dan OpenFace (GorillaFACS belum tersedia selama penelitian saat ini; Correia-Caeiro et al. 2025 ), aktivasi AU tertentu yang konsisten dan kesamaan dengan temuan dari spesies primata lain (Parr et al. 2007 ; Vick et al. 2007 ; Waller et al. 2015 ; Masai et al. 2022 ) mendukung hipotesis bahwa morfologi PF dan FPF telah dilestarikan di seluruh primata. Namun, PF dan FPF tidak sepenuhnya tumpang tindih secara morfologis dan tampaknya memiliki fungsi yang berbeda. Secara khusus, PF dapat meningkatkan variabilitas bermain, sementara FPF dapat meningkatkan persaingan selama sesi bermain dengan meningkatkan asimetri bermain. Dengan demikian, kami berhipotesis bahwa pada gorila, PF dan FPF dapat dipertahankan sebagai ekspresi yang berbeda karena mereka mungkin melayani tujuan adaptif yang berbeda. Hal ini selanjutnya didukung oleh pengamatan bahwa RFM mereka tampaknya berfungsi secara berbeda. Sementara FPF_RFM secara independen berkontribusi untuk memperpanjang sesi bermain dibandingkan dengan PF atau FPF yang tidak ditiru, PF_RFM mungkin memerlukan efek yang diperkuat dari FPF_RFM untuk mencapai hasil yang sama. Singkatnya, temuan kami dapat meletakkan dasar untuk penelitian lebih lanjut untuk mengonfirmasi atau mengungkap perbedaan fungsional dan/atau morfologis baru antara ekspresi wajah yang menyenangkan pada primata manusia dan non-manusia, menawarkan pemahaman yang lebih dalam tentang signifikansi adaptif dari mempertahankan dua ekspresi wajah dengan intensitas yang berbeda-beda sepanjang evolusi Hominidae.