
Abstrak
Sejarah ekologi dan perkembangan wilayah Chicago, Illinois, telah memengaruhi distribusi hutan saat ini di wilayah tersebut. Faktor-faktor yang sama ini, bersama dengan segregasi rasial yang sistemik dan berlangsung lama, telah membentuk distribusi populasi perkotaan yang mendapatkan manfaat dari layanan ekosistem yang disediakan oleh hutan perkotaan. Studi ini menunjukkan bahwa sejarah petak hutan terkait dengan atribut hutan seperti komposisi spesies pohon, kerapatan pohon, tinggi tajuk, dan heterogenitas struktural—yang semuanya merupakan prediktor penting dari kemampuan hutan untuk menyediakan layanan ekosistem. Namun, efek sejarah hutan ini hanya terlihat di inti hutan, karena tepi hutan serupa terlepas dari sejarah petak. Kami juga menemukan bahwa hutan di komunitas minoritas cenderung kurang mampu mendukung layanan ekosistem tingkat tinggi. Penelitian ini menunjukkan bahwa, ketika meningkatkan ekuitas hijau, penting untuk mempertimbangkan kapasitas variabel hutan untuk menyediakan layanan ekosistem.
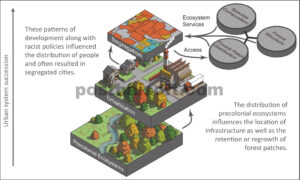
Distribusi spasial kelompok sosiodemografi dan ruang hijau di daerah perkotaan dibentuk oleh sejarah ekologi dan pembangunan kota. Sejarah ini telah mengakibatkan pola ketidakadilan ekologis, dengan komunitas yang kekurangan sumber daya dan/atau minoritas sering kali memiliki akses yang lebih sedikit ke ruang hijau (Schell et al . 2020 ). Kota-kota di Amerika Utara dibangun di atas tanah Pribumi, dan lanskapnya diubah seiring waktu menjadi jalan, saluran air, bangunan, tempat tinggal, dan ruang hijau yang membentuk ruang perkotaan saat ini (Roman et al . 2018 ; Ogden et al . 2019 ). Sepanjang perkembangan ini, kelompok ras dan etnis menetap di daerah yang berbeda. Sementara orang kulit putih yang kaya secara luas dapat memilih di mana mereka ingin tinggal, segregasi de jure dan eksplisit melarang orang-orang minoritas pindah ke lingkungan dengan berbagai fasilitas yang lebih luas, termasuk ruang hijau (Rothstein 2017 ). Bahasa Indonesia: Pada tahun 1935, Home Owners’ Loan Corporation, sebuah organisasi pemerintah yang dirancang untuk meningkatkan kepemilikan rumah setelah Depresi Besar, membuat peta keamanan pinjaman yang mengidentifikasi lingkungan yang dapat dan tidak dapat menerima pinjaman yang disetujui pemerintah. Proses ini, yang secara umum dikenal sebagai redlining, secara eksplisit berbasis ras, dan mengarahkan pengeluaran menjauh dari komunitas minoritas, dengan demikian semakin memperkuat segregasi di kota-kota AS (Rothstein 2017 ). Meskipun redlining secara resmi diakhiri dengan disahkannya Undang-Undang Perumahan yang Adil tahun 1968, konsekuensi dari kebijakan ini tetap ada hingga saat ini (Rothstein 2017 ). Populasi minoritas sering kali tinggal di komunitas dengan lebih banyak permukaan bangunan dan lebih sedikit pohon, membuat mereka lebih rentan terhadap kerugian lingkungan seperti efek pulau panas perkotaan (Hoffman et al . 2020 ) dan lebih kecil kemungkinannya untuk mendapatkan manfaat dari layanan ekosistem yang disediakan oleh ruang hijau (Schell et al . 2020 ).
Distribusi petak hutan juga telah dipengaruhi oleh perubahan historis dalam tutupan lahan dan transisi sosial ekonomi. Di sini, kami mendefinisikan petak hutan sebagai tempat (i) yang memiliki tajuk pohon terus menerus dan (ii) tempat pohon dapat beregenerasi secara spontan (Johnson et al . 2020 ). Petak hutan ini dapat terwujud dalam banyak cara: dari area alami skala besar yang analog dengan hutan yang lebih pedesaan hingga pengelompokan tajuk pohon kecil dan spontan yang tidak terkelola yang mungkin terjadi pada properti kosong atau di sepanjang jalur transit. Di Midwest AS, penebangan hutan untuk industri dan pertanian mencapai puncaknya pada akhir tahun 1930-an (Rhemtulla et al . 2007 ). Beberapa hutan terhindar dari pembukaan lahan tersebut dan tetap ada hingga saat ini, dan sejak saat itu, hutan baru kadang-kadang tumbuh kembali di tempat hutan asli pernah berdiri atau bahkan terbentuk di lahan yang sebelumnya tidak berhutan (misalnya padang rumput atau lahan basah). Meskipun waktunya berbeda, pola retensi, penebangan, dan pertumbuhan kembali hutan ini telah terjadi di seluruh Amerika Utara (misalnya Ogden et al . 2019 ).
Meskipun bagaimana hutan berubah seiring waktu dengan atau tanpa gangguan atau pengelolaan telah dipelajari secara luas, hingga saat ini daerah perkotaan sebagian besar dikecualikan dari penyelidikan ini. Dibandingkan dengan hutan di daerah pedesaan, hutan di kota mungkin memiliki pola suksesi yang sangat berbeda. Tanah perkotaan telah diubah oleh konstruksi, pengendapan polusi, dan limpasan pupuk (Pavao-Zuckerman 2008 ). Kota-kota, sebagai pusat perdagangan global, sering kali menjadi episentrum pengenalan spesies invasif (Von Der Lippe dan Kowarik 2008 ). Semua faktor tersebut telah menghasilkan rezim gangguan baru dan kumpulan spesies di hutan perkotaan (Johnson dan Handel 2016 ). Misalnya, di petak hutan di New York City, sebagian besar pohon tajuk adalah spesies asli tetapi hampir setengah dari pohon lantai hutan adalah spesies non-asli (Pregitzer et al . 2019 ).
Struktur, komposisi, dan distribusi hutan terkait dengan kemampuannya untuk menyediakan layanan ekosistem. Kanopi hutan yang lebih heterogen (yaitu yang memiliki tinggi kanopi dan celah vertikal yang bervariasi) memiliki lebih banyak ruang relung ekologis dan dapat menampung keanekaragaman hayati yang lebih besar (LaRue et al . 2019 ). Biomassa hutan berkorelasi positif dengan kemampuan hutan untuk menyimpan karbon, mengurangi limpasan air hujan, dan mengurangi efek pulau panas perkotaan (Nowak et al . 2008 ). Spesies invasif mengurangi kekayaan spesies tanaman dan sering kali menyediakan habitat berkualitas rendah untuk satwa liar asli (Knight et al . 2007 ), dan spesies pohon asli tertentu dapat mendukung lebih banyak keanekaragaman hayati (Fralish 2004 ). Banyak layanan ekosistem yang disediakan hutan dialami dalam jarak dekat, yang berarti bahwa orang yang tinggal di dekat hutan mendapat manfaat paling banyak. Ini terutama berlaku untuk layanan ekosistem budaya, seperti pengurangan stres, yang mengharuskan orang untuk berinteraksi secara fisik dengan hutan (Wolf et al . 2020 ). Mengatur layanan ekosistem, seperti pengurangan suhu dan mitigasi air hujan, tidak mengharuskan manusia berinteraksi dengan hutan, tetapi masih dialami di dekat batas hutan (Ziter et al . 2019 ).
Struktur dan komposisi tidak konsisten dalam petak-petak hutan. Tepi hutan lebih cerah, lebih berangin, lebih panas, dan lebih kering, dan lebih mungkin menghadapi polutan dari limpasan perkotaan atau pertanian (Matlack 1993 ). Karena itu, tepi hutan sering kali menampilkan lebih banyak spesies invasif dan pohon berukuran lebih kecil. Tepi hutan dapat memiliki efek beragam pada distribusi layanan ekosistem; di hutan beriklim sedang, tepi hutan sering kali menyimpan lebih banyak karbon di atas tanah daripada inti hutan (Smith et al . 2018 ). Orang-orang mungkin berinteraksi lebih langsung dengan tepi hutan dan dengan demikian mengalami lebih banyak layanan ekosistem budaya (Ziter et al . 2014 ), tetapi banyak spesies hewan sensitif tidak dapat tumbuh subur di tepi hutan (Hoover et al . 2006 ).
Di sini, kami menunjukkan dampak yang dimiliki warisan pembangunan perkotaan terhadap hutan dan komunitas manusia yang mendapatkan manfaat darinya (Gambar 1 ). Kami menggunakan peta hutan saat ini dan historis untuk mengidentifikasi perubahan dalam luasan petak hutan selama dua abad terakhir, kemudian mengeksplorasi bagaimana sejarah itu memengaruhi karakteristik hutan saat ini melalui pengambilan sampel di lapangan dan analisis struktural lidar (deteksi cahaya dan pengukuran jarak). Akhirnya, kami menunjukkan bagaimana distribusi petak hutan dengan tipe sejarah yang berbeda (dan kapasitas yang berbeda untuk menawarkan layanan ekosistem) bervariasi dengan distribusi ras komunitas. Kami berhipotesis bahwa: (i) sejarah retensi, kerusakan, dan/atau pertumbuhan kembali hutan memengaruhi komposisi dan struktur spesies pohon saat ini; (ii) pengaruh historis ini memengaruhi inti dan tepi hutan secara berbeda; dan (iii) ada disparitas ras dalam kedekatan penduduk dengan tutupan hutan dengan sejarah yang berbeda.
Metode
Proyek kami difokuskan pada petak-petak hutan di wilayah tujuh kabupaten Chicago, Illinois (luas: 1,05 juta ha, populasi: 9,7 juta). Kota Chicago (populasi: 2,7 juta) didirikan pada tahun 1837 dan merupakan kota terbesar ketiga di AS. Wilayah ini beragam; misalnya, pada tahun 2015, 23% populasi wilayah tersebut adalah orang kulit hitam, 54% orang kulit putih, dan 25% adalah Hispanik (US Census Bureau 2023 ). Wilayah ini terletak di persimpangan padang rumput dan hutan berdaun lebar gugur bagian timur. Sebelum kolonisasi Euro-Amerika, wilayah ini merupakan rumah bagi Three Fires Confederacy (Native Land Digital 2020 ). Di bawah perawatan mereka, wilayah tersebut 60–80% berupa padang rumput. Hutan terbatas pada daerah yang terlindungi dari kebakaran rutin yang digunakan suku-suku ini untuk mengelola lanskap, dan hutan-hutan ini memiliki kanopi yang relatif terbuka dan didominasi oleh pohon ek ( Quercus spp; McBride dan Bowles 2001 ).
Untuk mengukur riwayat gangguan petak hutan, kami menggunakan lapisan petak hutan dari tiga periode waktu: 2010 (Darling et al . 2023 ), 1939 (Fahey dan Casali 2017 ), dan 1830 (McBride dan Halsey 2015 ). Kami mengklasifikasikan riwayat petak dengan menilai tumpang tindih petak saat ini dengan lapisan 1939 dan 1830. Jenis riwayat ditetapkan sebagai sisa (berhutan di ketiga periode waktu), pertumbuhan kembali (berhutan pada 2010 dan 1830 tetapi tidak pada 1939), dan baru (berhutan saat ini tetapi tidak berhutan pada era prakolonial) (Lampiran S1 : Gambar S1a ). Tepi dan inti petak diidentifikasi oleh Darling et al . ( 2023 ), dengan area tepi memanjang 15 m ke dalam petak. Meskipun beberapa efek tepi meluas lebih dalam ke petak hutan, di hutan beriklim sedang perbedaan komposisi dan struktur spesies umumnya terjadi dalam jarak 20 m dari tepi hutan (Franklin et al . 2021 ).
Bahasa Indonesia: Kami mengukur komposisi spesies saat ini dan perbedaan struktural di seluruh riwayat petak hutan pada musim panas tahun 2020 dan 2021 dengan mengambil sampel dua puluh plot 0,4 ha di area inti dan tepi setiap jenis riwayat petak—baru, tumbuh kembali, dan sisa (total 120 plot) (Lampiran S1 : Gambar S1b ). Di setiap plot, kami mencatat spesies dan diameter setinggi dada (DBH) setiap pohon dengan DBH >2,54 cm, lalu menghitung luas basal (BA) setiap spesies dalam setiap plot. Semua analisis statistik dan visualisasi dilakukan dalam R (v4.3.0; Tim Inti R 2023 ). Perbedaan komposisi spesies plot divisualisasikan dengan ordinasi penskalaan multidimensi non-metrik (NMDS), yang dijalankan di tiga dimensi dan dengan 10.000 iterasi. Kami kemudian menggunakan fungsi envfit dari paket vegan (yang menyesuaikan faktor lingkungan dengan ordinasi dan menyediakan nilai kesesuaian dan signifikansi) dalam R untuk menguji apakah jenis riwayat hutan menjelaskan perbedaan dalam komposisi plot (Oksanen et al . 2022 ). Kami juga membandingkan perbedaan dalam total BA dan kepadatan batang di seluruh riwayat petak dengan analisis varians satu arah (ANOVA), tanpa menyertakan efek acak atau kovariat.
Kami mengukur perbedaan dalam struktur hutan menggunakan lidar udara yang diperoleh dari Survei Geospasial Negara Bagian Illinois ( 2023 ). Titik-titik lidar ini sudah diklasifikasikan, termasuk nilai pohon, tanah, dan bangunan. Titik-titik tersebut dipotong ke luasan plot, dan kebisingan serta tinggi tajuk yang dinormalisasi disaring dengan mengurangi tinggi tanah dari setiap titik tajuk. Selanjutnya, kami menghitung kompleksitas eksterior (kekasaran puncak), kompleksitas interior (deviasi standar vertikal), kerapatan titik, dan tinggi tajuk maksimum (maks) rata-rata, mengikuti LaRue et al . ( 2019 ). Singkatnya, kompleksitas eksterior adalah heterogenitas atau tonjolan di bagian atas tajuk; tajuk yang dibentuk oleh pohon-pohon yang lebih pendek dan lebih tinggi yang diselingi akan memiliki kompleksitas eksterior yang lebih tinggi. Kompleksitas interior adalah heterogenitas di dalam tajuk; hutan yang memiliki beberapa lapisan atau lubang di dalam tajuknya akan memiliki kompleksitas interior yang tinggi. Kerapatan titik adalah jumlah titik tajuk per meter persegi—hutan dengan cabang yang lebih padat akan memiliki kerapatan titik yang lebih tinggi. Terakhir, tinggi tajuk maksimum rata-rata adalah tinggi rata-rata bagian atas tajuk. Ilustrasi perbedaan ini tersedia di Lampiran S1 : Gambar S2 . Kami membandingkan nilai-nilai struktural ini di seluruh riwayat patch menggunakan ANOVA satu arah, sekali lagi tanpa menyertakan efek acak atau kovariat.
Kami membandingkan pola dalam distribusi petak hutan dan susunan ras menggunakan data skala blok-kelompok dari rata-rata lima tahun Survei Komunitas Amerika 2015 (Kantor Sensus AS 2023 ). Blok-kelompok adalah skala terbaik yang digunakan Biro Sensus AS untuk menggabungkan banyak data sosiodemografi, tetapi ukurannya bervariasi karena dibuat untuk memastikan bahwa antara 600 dan 3000 orang berada di dalam batas-batasnya. Oleh karena itu, kami menggabungkan data tersebut ke dalam kisi 2 km menggunakan rata-rata tertimbang area (Lampiran S1 : Gambar S1c ). Banyak variabel terkait ras, pendidikan, dan kekayaan berkorelasi di Amerika Utara karena rasisme sistemik (Rothstein 2017 ). Kami berfokus pada satu ukuran status sosiodemografi (persentase populasi yang berkulit putih) untuk menyederhanakan analisis ini. Kami juga menghitung luas riwayat petak hutan dan tepi serta inti di kisi yang sama. Kami memodelkan hubungan antara populasi kulit putih dan tutupan hutan menggunakan model gamma rintangan Bayesian dari paket brms di R karena tutupan petak hutan digelembungkan ke nol dan didistribusikan gamma (Bürkner 2017 ). Model, yang dijalankan melintasi empat rantai dengan 5000 iterasi dan pemanasan 500 iterasi, menilai hubungan antara ras dan tutupan hutan dengan dua cara: dengan meregresikan persentase populasi kulit putih ke area setiap tipe riwayat hutan dan dengan menghitung probabilitas bahwa sel grid akan memiliki area hutan nol. Kami menggunakan metode yang sama untuk membandingkan area tepi dan inti di setiap sel grid dengan susunan ras.
Hasil
Petak-petak hutan menutupi lebih dari 73.000 ha (7%) wilayah Chicago. Petak-petak hutan yang tersisa rata-rata lebih besar (rata-rata = 1,1 ha), masing-masing sekitar tiga kali dan dua kali ukuran petak-petak hutan yang tumbuh kembali dan petak-petak hutan baru. Petak-petak hutan di wilayah tersebut sangat terfragmentasi, tercermin dalam temuan bahwa total luas petak dibagi secara merata antara inti dan tepi. Namun, petak-petak hutan yang tersisa kurang terfragmentasi, dengan 62% luasnya berada di dalam inti: kira-kira 20% lebih banyak daripada di petak-petak hutan baru dan yang tumbuh kembali (Lampiran S1 : Tabel S1 ).
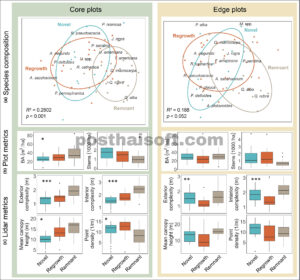
Komposisi spesies di area hutan inti bervariasi di antara kategori riwayat petak (Gambar 2a ; Lampiran S1 : Tabel S2 ). Plot sisa di area inti memiliki dominasi pohon ek yang lebih tinggi (63% dari total BA), empat kali lipat dari plot pertumbuhan kembali dan 15 kali lipat dari plot baru. Spesies invasif terdiri dari 21% BA di plot inti baru—lima kali lipat dari plot sisa dan lebih dari dua kali lipat dari plot pertumbuhan kembali. European buckthorn ( Rhamnus cathartica ) sendiri terdiri dari 16% BA di plot baru. Plot inti pertumbuhan kembali memiliki beberapa pohon ek, yang mirip dengan plot inti sisa, tetapi juga memiliki lebih banyak penduduk asli yang toleran terhadap naungan (misalnya box elder [ Acer negundo ] dan black walnut [ Juglans nigra ]) dan proporsi invasif yang lebih tinggi daripada plot sisa. Kami tidak menemukan perbedaan signifikan dalam komposisi spesies di tepi hutan menurut tipe riwayat petak. Di semua wilayah, spesies invasif mencakup 13% BA, dengan buckthorn Eropa sebagai penyumbang paling banyak (8% dari total BA).
Gambar 2
Komposisi dan struktur spesies berbeda di tiga riwayat petak—baru, tumbuh kembali, dan sisa—di plot inti (kiri) tetapi tidak di plot tepi (kanan). (a) Komposisi spesies di seluruh riwayat petak didefinisikan menggunakan ordinasi penskalaan multidimensi non-metrik (NMDS). (b dan c) Perbedaan struktural diidentifikasi menggunakan analisis varians (ANOVA) pada dua metrik berbasis plot dan empat metrik berbasis lidar udara (deteksi cahaya dan pengukuran jarak). Signifikansi statistik ditunjukkan sebagai berikut: * P < 0,05, ** P < 0,01, dan *** P < 0,001. Garis horizontal di dalam kotak menggambarkan nilai median, kotak mewakili rentang interkuartil (persentil ke-25–75), kumis (garis vertikal) mewakili rentang interkuartil 1,5×, dan lingkaran padat menggambarkan outlier.
Plot inti sisa memiliki pohon yang lebih sedikit tetapi lebih besar daripada plot inti baru, dengan setengah batang tetapi 20% BA lebih tinggi, sementara plot inti pertumbuhan kembali memiliki nilai menengah (Gambar 2b ). Perbedaan struktural ini juga terlihat dari analisis lidar udara. Plot inti sisa memiliki kompleksitas interior dan eksterior yang lebih tinggi dan kepadatan titik yang lebih rendah daripada plot inti baru dan pertumbuhan kembali (Gambar 2c ). Plot inti sisa juga memiliki tinggi tajuk rata-rata yang lebih tinggi (18,8 m), sedangkan plot baru memiliki yang terendah (12,5 m). Di plot tepi, tidak ada perbedaan signifikan dalam kepadatan batang, BA, dan titik di seluruh riwayat petak. BA rata-rata di plot tepi (27,7 m 2 /ha) mirip dengan BA rata-rata di plot inti baru (26,1 m 2 /ha). Di antara riwayat petak di plot tepi, meskipun tinggi tajuk rata-rata tidak berbeda secara signifikan, ada perbedaan dalam kompleksitas. Tepi sisa dan tepi pertumbuhan kembali adalah yang paling kompleks dan paling tidak kompleks, berturut-turut. Untuk pemahaman yang lebih komprehensif tentang struktur hutan, sebaiknya bandingkan serangkaian metrik yang diperoleh dari lidar. Lihat Lampiran S1 : Gambar S3 untuk metrik tambahan dari analisis ini.
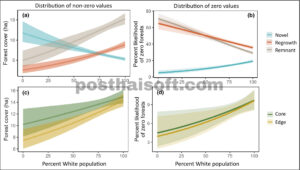
Di wilayah Chicago, daerah dengan proporsi penduduk kulit putih yang lebih tinggi cenderung memiliki cakupan hutan (yang sebagian besar merupakan sisa) yang lebih besar (Gambar 3 ; Lampiran S1 : Tabel S3 ). Karena datanya digelembungkan ke nol, kami membandingkan data petak persentase penduduk kulit putih dengan area setiap tipe hutan (Gambar 3a ) dan juga dengan kemungkinan tidak ada area hutan (Gambar 3b ). Kami menemukan bahwa daerah dengan persentase penduduk kulit putih terendah memiliki cakupan area hutan baru yang lebih besar, dan cakupan area hutan tumbuh kembali dan sisa yang lebih kecil (Gambar 3a ). Demikian pula, daerah dengan persentase penduduk kulit putih terendah cenderung tidak memiliki hutan sisa atau hutan tumbuh kembali, tetapi hampir selalu memiliki beberapa hutan baru (Gambar 3b ). Sebaliknya, di daerah dengan persentase penduduk kulit putih tertinggi, sekitar 30% tidak memiliki hutan sisa atau hutan tumbuh kembali di dekatnya dan 20% tidak memiliki hutan baru. Proporsi inti dan tepi lebih merata, meskipun area dengan penduduk kulit putih paling sedikit memiliki rasio inti-tepi yang sedikit lebih tinggi (Gambar 3 , c dan d).
Gambar 3
Distribusi kelompok ras dan petak hutan dengan sejarah yang berbeda tidak seragam di seluruh wilayah Chicago. Daerah dengan persentase penduduk kulit putih yang lebih tinggi memiliki cakupan hutan yang lebih luas, didominasi oleh hutan sisa (atas); namun, rasio tepi dan inti relatif merata di seluruh gradien ras (bawah). Arsir menunjukkan interval kepercayaan 95%.
Diskusi
Berabad-abad pembangunan dan pengelolaan Euro-Amerika mengubah komposisi dan struktur hutan di wilayah Chicago, dan perubahan ini terkait dengan kapasitas hutan untuk menyediakan layanan ekosistem. Hutan dengan biomassa lebih tinggi, tajuk heterogen secara struktural, lebih sedikit spesies invasif, dan lebih banyak spesies kunci seperti pohon ek dapat menyediakan lebih banyak layanan ekosistem. Kompleksitas struktural yang diperoleh dari Lidar dan pengukuran struktur dan komposisi hutan di lapangan menghasilkan gambaran yang konsisten: inti hutan sisa memiliki potensi lebih besar untuk menyediakan layanan ekosistem daripada inti hutan baru dan hutan yang tumbuh kembali. Inti hutan sisa menyerupai hutan prakolonial, karena memiliki proporsi pohon ek yang lebih tinggi, lebih sedikit pohon secara keseluruhan, lebih sedikit spesies invasif, dan lebih banyak tajuk terbuka. Namun, inti hutan yang tumbuh kembali dan baru memiliki atribut yang berlawanan: kepadatan pohon lebih tinggi, biomassa lebih rendah, dan lebih banyak spesies invasif dan toleran naungan.
Studi ini menyoroti pentingnya melindungi hutan sisa untuk melestarikan keanekaragaman hayati asli. Pohon ek yang mendominasi inti hutan sisa adalah spesies kunci di hutan midwest, dan mereka mendukung banyak flora dan fauna di wilayah tersebut (Fralish 2004 ). Sebagian besar hutan prakolonial di wilayah tersebut telah hilang (Fahey dan Casali 2017 ), dan melindungi apa yang tersisa sangat penting. Banyak dari petak hutan yang tumbuh kembali telah tumbuh selama 80 tahun, namun mereka belum mendapatkan kembali karakteristik struktural (heterogenitas tinggi) dan komposisi (didominasi pohon ek) yang mendukung keanekaragaman hayati dari hutan sisa. Mengingat banyaknya spesies invasif dan toleran naungan di petak yang tumbuh kembali dan baru, hutan-hutan ini tidak mungkin kembali ke ekosistem pohon ek kompleks yang pernah menjadi ciri wilayah tersebut tanpa intervensi aktif (Nowacki dan Abrams 2008 ).
Meskipun struktur dan komposisi spesies mereka saat ini menyerupai kondisi prakolonial, hutan sisa mungkin memerlukan pengelolaan aktif untuk mempertahankan karakteristik ini. Hutan prakolonial di wilayah Chicago berkembang sebagai hasil dari perawatan Pribumi, yang mencakup penggunaan api intensitas rendah (Pyne 1982 ); namun, seiring berkembangnya wilayah tersebut, kebakaran sebagian besar ditekan. Spesies pohon ek dalam penelitian ini (terutama pohon ek bur [ Quercus macrocarpa ] dan pohon ek putih [ Quercus alba ]) tidak tahan naungan, dan untuk mempertahankan hutan heterogen yang didominasi pohon ek di masa depan, perlu dibuat bukaan kanopi yang memungkinkan cahaya mencapai lantai hutan dan memulihkan rezim kebakaran untuk mempertahankan kondisi terbuka ini (Nowacki dan Abrams 2008 ). Hutan perkotaan menghadapi tantangan tambahan untuk mempertahankan keanekaragaman hayati yang tinggi; hutan perkotaan sering kali menjadi episentrum untuk pengenalan spesies invasif, dan limpasan pupuk serta polusi dapat merusak tanah hutan (Johnson dan Handel 2016 ). Manajemen intensif jangka panjang mungkin diperlukan untuk menghilangkan spesies invasif dari hutan-hutan ini (Johnson dan Handel 2019 ). Selain itu, hutan-hutan dalam studi ini sangat terfragmentasi dan diselingi dengan pembangunan. Pembakaran yang direncanakan umumnya digunakan untuk membatasi pertumbuhan spesies yang tidak tahan api dan invasif di ekosistem yang didominasi pohon ek, tetapi di daerah perkotaan, kehati-hatian harus dilakukan untuk memastikan bahwa kebakaran dan asap tidak memiliki efek buruk pada orang atau properti (Hu et al . 2008 ). Pertumbuhan kembali dan hutan baru dapat memperoleh manfaat dari teknik-teknik manajemen yang sama ini tetapi kemungkinan akan memerlukan upaya yang lebih ekstensif, karena hutan-hutan ini saat ini memiliki lebih banyak spesies invasif dan—mengingat sejarah perkembangannya—kemungkinan memiliki tanah yang lebih terganggu.
Sementara riwayat petak merupakan prediktor yang baik untuk komposisi dan struktur di inti hutan, tepiannya serupa terlepas dari riwayat petak. Tepi hutan secara konsisten memiliki kelimpahan spesies invasif yang lebih tinggi dan BA yang lebih rendah daripada plot inti. Tepi hutan cenderung mengalami suhu, angin, dan tingkat paparan polusi yang lebih tinggi (Matlack 1993 ), dan faktor-faktor ini tampaknya telah mengalahkan warisan historis lintasan pengembangan petak hutan, membuat struktur dan komposisinya serupa dengan inti baru. Pengembangan masa lalu memecah-belah hutan di wilayah Chicago; hampir setengah dari semua kawasan hutan berada dalam jarak 15 m dari tepi petak (Lampiran S1 : Tabel S1 ). Pengembangan di masa depan yang mendorong fragmentasi akan mengurangi rasio inti-ke-tepi petak hutan dan terus menurunkan manfaat yang terkait dengan inti hutan. Kebijakan yang ditujukan untuk membatasi kerusakan hutan dapat mencegah fragmentasi lebih lanjut dan melindungi hutan inti yang beragam hayati dan menyediakan pasokan tinggi.
Banyak upaya untuk meningkatkan ekuitas hijau hanya bergantung pada identifikasi kesenjangan dalam luas tajuk pohon, tetapi gagal memperhitungkan fakta bahwa petak-petak hutan dapat memiliki kapasitas yang berbeda untuk menyediakan layanan ekosistem. Hutan baru sering kali merupakan satu-satunya hutan yang ditemukan dalam komunitas kulit berwarna, dan kemungkinan besar menyediakan lebih sedikit layanan ekosistem penyediaan—seperti mengurangi efek pulau panas perkotaan dan limpasan air hujan—daripada hutan sisa. Selain itu, beberapa manfaat kesehatan manusia yang terkait dengan hutan perkotaan, seperti mengurangi stres dan tekanan darah, mengharuskan orang untuk berinteraksi langsung dengan mereka (Wolf et al . 2020 ). Hutan baru di Chicago memiliki banyak pohon kecil, dicirikan oleh tajuk yang lebih rendah dan lebih rapat, dan secara visual berbeda dari hutan yang tumbuh kembali dan hutan sisa (Lampiran S1 : Gambar S1 ). Faktor-faktor ini selanjutnya dapat menghalangi anggota masyarakat lokal untuk mengunjungi petak-petak ini dan mengalami manfaat mental dan fisik yang terkait dengan interaksi dengan hutan, kemungkinan yang memerlukan studi lebih lanjut. Kenyataannya, hutan-hutan ini mungkin lebih merupakan suatu beban daripada suatu aset, mengingat bahwa hutan-hutan tersebut seringkali tumbuh di lahan-lahan kosong, seringkali menjadi indikator divestasi dan kurangnya pengelolaan, dan mungkin digunakan untuk pembuangan atau untuk menyembunyikan kegiatan-kegiatan terlarang (Riley et al . 2018 ).
Pengembangan perkotaan masa depan yang memprioritaskan perlindungan dan pengelolaan hutan berkualitas tinggi, memulihkan hutan berkualitas rendah, dan menanam hutan atau ruang hijau baru di komunitas minoritas dapat meningkatkan akses yang adil terhadap keanekaragaman hayati dan layanan ekosistem. Namun, kegiatan restorasi dan pengelolaan ini memerlukan dukungan masyarakat. Komunitas sosial ekonomi yang beragam mungkin memiliki preferensi yang berbeda untuk ruang hijau dan hutan perkotaan (Ordóñez-Barona 2017 ). Studi ini menggunakan persentase populasi kulit putih untuk mengidentifikasi tren dalam akses yang tidak adil ke hutan berkualitas tinggi, tetapi ada keragaman yang luar biasa di seluruh spektrum sosial ekonomi ini. Mengenali keragaman ini sangat penting tidak hanya untuk mengembangkan pemahaman yang lebih baik tentang ketidakadilan, tetapi juga untuk menanggapinya. Berkomunikasi dengan komunitas yang beragam sangat penting untuk memastikan bahwa upaya untuk meningkatkan ekuitas hijau mencerminkan preferensi komunitas dan memungkinkan penduduk untuk mencapai tujuan masa depan mereka.
Ucapan Terima Kasih
Penulis ingin menyampaikan penghargaan yang sebesar-besarnya atas kontribusi dari Dexter Locke, John Paul Schmit, Michelle Johnson, dan Nancy Sonti. Tulisan ini tidak akan mungkin terwujud tanpa mereka. Penulis mengucapkan terima kasih kepada Brett Jelinek dan Ian Schuman atas bantuan mereka dalam kerja lapangan. Penelitian ini didukung oleh beasiswa dari Pusat Ilmu Pohon Morton Arboretum dan Pusat Sintesis Sosial-Lingkungan Nasional (SESYNC) dengan pendanaan yang diterima dari Yayasan Sains Nasional DBI-1639145.