
Abstrak
Meramalkan manfaat intervensi manajemen yang dimaksudkan untuk meningkatkan kondisi ekologis memerlukan pemahaman kausal tentang faktor-faktor yang menyebabkan perubahan sistem. Atribusi kausal suatu faktor didefinisikan sebagai perbedaan antara hasil yang diamati dengan adanya faktor dan hasil yang akan diamati jika faktor tersebut tidak ada, yaitu, kondisi kontrafaktual. Memperkirakan kontras ini relatif mudah, jika kontrol yang cocok atau acak tersedia untuk memperkirakan kondisi kontrafaktual. Namun, peneliti harus bernalar secara retrospektif dari data observasional jika kontrol yang cocok atau acak tidak tersedia. Dalam kasus ini, tantangan dalam menetapkan atribusi kausal adalah dalam memperkirakan kontrafaktual yang sebenarnya, yaitu, hasil yang akan dihasilkan dari ketidakhadiran faktor tersebut, mengingat faktor tersebut ada. Analisis kausal memungkinkan estimasi kontrafaktual dari data observasional, dengan asumsi bahwa model menangkap semua penyebab umum antara paparan dan hasil, bahwa paparan tidak tergantung pada faktor lain dalam model (yaitu, eksogen), dan bahwa paparan menyebabkan perubahan arah yang sama untuk semua unit (yaitu, monotonik). Saya memperkirakan secara retrospektif atribusi kausal faktor-faktor terkait habitat terhadap tingkat rekrutmen pada populasi boreal rusa kutub hutan Kanada ( Rangifer tarandus caribou ). Gangguan habitat agregat memiliki atribusi kausal yang rendah (17,6%). Atribusi lebih besar (29,5%) ketika gangguan habitat dipecah menjadi faktor-faktor berbeda yang terkait dengan jalur penurunan rusa kutub yang berbeda. Atribusi kausal semua faktor habitat yang dipertimbangkan meskipun demikian jarang melebihi 50%, menunjukkan bahwa ada faktor sistematis dan/atau stokastik lain yang dapat membatasi efektivitas tindakan pemulihan terkait habitat saat ini. Diperlukan upaya lebih besar untuk memahami faktor-faktor ini dan bagaimana mereka dapat dikelola untuk meningkatkan kemungkinan pemulihan rusa kutub yang berhasil.
PERKENALAN
Tantangan utama dalam konservasi adalah pengelolaan sistem kompleks yang menghadapi banyak faktor yang saling berinteraksi (Côté et al., 2016 ). Memahami efek relatif dari faktor-faktor yang menyebabkan degradasi sistem sangat penting untuk merancang intervensi pengelolaan untuk meningkatkan kondisinya (Hobbs & Norton, 1996 ). Atribusi kausal suatu faktor didefinisikan sebagai perbedaan antara hasil yang diamati dengan adanya faktor dan hasil yang akan diamati tanpa adanya faktor (Yamamoto, 2012 ). Hasil terakhir ini disebut sebagai kontrafaktual dan dapat diperkirakan dalam pengaturan eksperimental menggunakan kontrol acak atau yang dicocokkan (Coetzee & Gaston, 2021 ; Ferraro, 2009 ). Pengamatan semacam itu hanya memperkirakan kondisi kontrafaktual karena kontrafaktual yang sebenarnya tidak akan pernah dapat diamati; unit tertentu terpapar pada suatu faktor atau tidak, tetapi tidak keduanya (Dawid, 2000 ).
Sayangnya, bahkan kontrafaktual yang didekati pun tidak layak dilakukan ketika kesimpulan tentang penyebab harus ditarik secara retrospektif hanya dari data observasi. Misalnya, kita sering ingin menentukan penyebab penurunan spesies setelah itu terjadi, dan bukti mungkin sebagian besar dibatasi pada rangkaian waktu estimasi populasi dan perubahan keadaan faktor-faktor yang mungkin berkontribusi. Paling umum, metode pemilihan model korelatif diterapkan untuk memprediksi tren populasi menggunakan set faktor kandidat, dan set kovariat ini yang menjelaskan varians paling banyak umumnya diasumsikan telah menyebabkan tren yang diamati (Arif & MacNeil, 2022 ; Burnham & Anderson, 2011 ). Tetapi sementara model tersebut memiliki kekuatan penjelasan, tidak ada alasan apriori untuk berasumsi bahwa mereka secara akurat mewakili efek kausal (Addicott et al., 2022 ; Oliver & Roy, 2015 ).
Sementara analisis kausal (Pearl, 2009 ) semakin banyak diterapkan dalam ekologi (Kimmel et al., 2021 ; Larsen et al., 2019 ; Law et al., 2017 ; Wilson et al., 2021 ), penerapan penalaran kontrafaktual pada isu kritis atribusi kausal belum. Di sini, saya menerapkan analisis kausal kontrafaktual pada isu gangguan habitat dan dampaknya pada kinerja demografi rusa kutub hutan ( Rangifer tarandus caribou ) di antara subpopulasi boreal di Kanada. Saya menunjukkan bagaimana kontrafaktual dapat diterapkan untuk memperkirakan atribusi kausal faktor-faktor, di bawah asumsi yang kuat, ketika hanya data retrospektif non-eksperimental yang tersedia.
METODE
Mengidentifikasi efek kausal dari data observasi

Perbedaan antara “mengamati” (Persamaan 1 ) dan “melakukan” (Persamaan 2 ) merupakan perbedaan utama dalam penalaran kausal dan membedakan dua anak tangga pertama dari “tangga kausalitas” Pearl ( 2019 ). Meskipun pengamatan
Dan
dapat terlihat terjadi bersamaan, hanya jika ada intervensi untuk mengubah nilai
perubahan
dapatkah kita mengklaim bahwa variabel-variabel tersebut berhubungan secara kausal. Beralih dari “mengamati” ke “melakukan” juga memerlukan informasi lebih lanjut tentang sistem yang sedang diselidiki. “Mengamati” suatu hubungan hanya memerlukan pengamatan yang cukup
Dan
untuk menyimpulkan bahwa koefisien korelasi mereka berbeda secara signifikan dari nol. Untuk memprediksi pengaruhnya terhadap
dari campur tangan pada
memerlukan bukti eksperimental atau, jika tidak ada, penerapan alat analisis kausal:
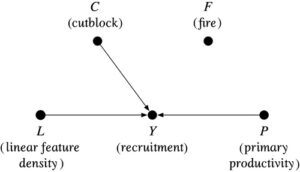
- Model kausal struktural (SCM), dinyatakan dalam kerangka Pearl ( 2009 ) sebagai grafik asiklik terarah (DAG), yang menentukan semua jalur kausal melalui busur terarah (yaitu, panah) antara paparanXdan hasilY, termasuk semua “penyebab umum” yang diamati dan tidak diamati antara paparan dan hasil (Gambar 1 ), serta fungsi yang mendefinisikan hubungan antara node yang dihubungkan oleh jalur.
- Seperangkat penyesuaian statistik yang memblokir semua jalur non-kausal tanpa memblokir semua jalur kausal, menurut serangkaian kriteria identifikasi kausal (yaitu, pengkondisian pada variabel untuk mencapai “pemisahan-d”) (Grace & Irvine, 2020 ; Pearl, 2009 ).
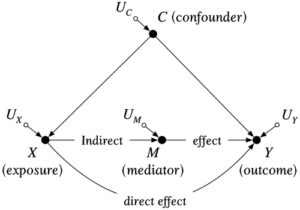
Contoh grafik asiklik terarah untuk merepresentasikan struktur kausal suatu sistem, menghubungkan variabel acak (“simpul”) melalui anak panah (“busur terarah”) untuk merepresentasikan hubungan kausal. Paparan menyebabkan suatu hasil melalui efek langsung atau secara tidak langsung melalui mediator . Faktor pengganggu adalah penyebab umum dari paparan dan hasil. Memperkirakan efek kausal dari pada dalam hal ini memerlukan penyesuaian statistik untuk memblokir jalur “pintu belakang” antara paparan dan hasil melalui faktor pengganggu Tidak berkorelasi Faktor-faktor tersebut mewakili variasi antar individu yang tidak dijelaskan oleh model.
Terkadang, tidak ada serangkaian penyesuaian yang memungkinkan untuk mengisolasi efek kausal
pada
Misalnya, suatu faktor pengganggu mungkin diketahui atau diduga tetapi tidak teramati, sehingga mencegah penyesuaiannya dan dengan demikian mencegah pemblokiran jalur non-kausal. Dalam kasus seperti itu, efek kausal yang tidak bias pada
dari intervensi
=
tidak dapat diperkirakan dari kumpulan data observasional. Misalnya, mengamati korelasi positif antara perburuan predator dan konflik yang berkaitan dengan manusia dengan predator pada tahun-tahun berikutnya menunjukkan bahwa perburuan menyebabkan peningkatan konflik (misalnya, Teichman et al., 2016 ); namun, tanpa penyesuaian yang memadai untuk ukuran populasi predator dari waktu ke waktu, yang jarang diketahui tetapi berhubungan secara kausal dengan jumlah pembunuhan oleh pemburu dan kejadian konflik, hubungan kausal yang tidak bias antara perburuan dan konflik tidak dapat disimpulkan.
Pearl ( 2009 ) memperluas metode analisis kausal untuk juga bernalar secara kontrafaktual; yaitu, untuk membuat inferensi retrospektif tentang bagaimana suatu hasil akan berbeda jika suatu paparan berbeda. Pernyataan “jika” menyajikan suatu kondisi hipotetis yang umumnya disebut anteseden (Pearl et al., 2016 ). Analisis kontrafaktual memperkirakan bagaimana suatu hasil akan berbeda jika hanya anteseden yang terjadi, dan semua faktor lainnya tetap konstan. Ini memberikan tingkat penalaran ketiga dan melengkapi “tangga sebab akibat” Pearl ( 2019 ) (Tabel 1 ). Kontrafaktual secara konseptual mirip dengan peran kelompok kontrol dalam uji coba terkontrol acak (RCT). Keduanya bertujuan untuk memberikan dasar untuk memahami apa yang akan terjadi jika tidak adanya paparan; Namun, kelompok kontrol dalam RCT memberikan perbandingan “dunia nyata” terhadap kelompok yang terpapar, sementara kontrafaktual, dalam logika Pearl ( 2009 ), adalah konsekuensi hipotetis dari tidak terpapar, seperti yang disimpulkan dari model kausal.
| Pertanyaan | Perwakilan | Pertanyaan | Aplikasi |
|---|---|---|---|
| “Mengamati” | P(Y|x) | Apa ituYketika kami mengamatiX=x…. | Menginterpolasi hasil dari hubungan korelatif dengan paparan |
| “Melakukan” (yaitu, melakukan intervensi) | P[Y|do(x)] | Apa yang akanYjadi jika kita ingin mengaturnyaX=x…. | Menerapkan pengobatan dan mengamati hasilnya |
| Penalaran kontrafaktual | P(Yx|x′,y′) | Apa yang akanYsudah pernah adaX=xdaripada pengobatan yang sebenarnyaX=x′, yang menghasilkan hasil yang diamatiY=y′ | Memperkirakan hasil yang akan terjadi jika pengobatan yang berbeda (atau tidak ada pengobatan) diterapkan |
Sebab akibat yang diperlukan dan cukup
Mengikuti Pearl ( 2022 ) dan menggunakan notasi Hannart et al. ( 2016 ), pertimbangkan kasus paparan biner terhadap faktor
dan hasil biner yang dihasilkan
Hasil “faktual” atau hasil dunia nyata didefinisikan sebagai:


Misalnya, pertimbangkan program pengurangan serigala yang diterapkan untuk meningkatkan laju pertumbuhan rusa kutub gunung selatan di wilayah pegunungan di Kanada bagian barat (Lamb et al., 2024 ). Dalam kasus ini, di antara sebagian wilayah pegunungan yang menerima penanganan, probabilitas kausalitas yang diperlukan (PN) menyatakan probabilitas bahwa laju pertumbuhan rusa kutub tidak akan meningkat jika pengurangan serigala tidak terjadi. Di antara sebagian wilayah pegunungan yang tidak menerima penanganan, probabilitas kausalitas yang cukup (PS) menyatakan probabilitas bahwa laju pertumbuhan rusa kutub akan meningkat jika pengurangan serigala terjadi. Menggabungkan kontrafaktual ini ke dalam probabilitas kausalitas yang diperlukan dan cukup menyatakan probabilitas bahwa peningkatan laju pertumbuhan rusa kutub akan terjadi dengan paparan penanganan pengurangan serigala dan tidak akan terjadi jika tidak ada penanganan.
Tian dan Pearl ( 2000 ) menunjukkan bahwa untuk melakukan penalaran kontrafaktual dari kuantitas-kuantitas ini hanya berdasarkan data observasional, diperlukan asumsi-asumsi tambahan untuk menghasilkan estimasi titik kausalitas. Pertama, paparan
harus “eksogen;” yaitu, tidak boleh ada busur masuk di SCM yang sesuai. Kedua, efek dari
harus “monotonik,” yang berarti bahwa hasil paparan untuk semua unit harus dalam arah yang sama (Manski, 1997 ). Jadi biasanya, suatu faktor harus diasumsikan selalu memiliki efek negatif di antara semua yang terpapar, dibandingkan memiliki efek perlindungan untuk beberapa. Jika asumsi ini berlaku, maka PN, PS, dan PNS dapat dihitung dari
Dan
(Persamaan 3 dan 4 ) sebagai berikut:
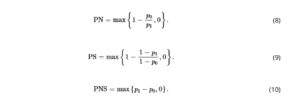
Atribusi kausal
Berdasarkan data observasi saja, kita sekarang memiliki langkah-langkah dan asumsi minimum yang diperlukan untuk memperkirakan atribusi suatu hasil terhadap penyebab tertentu. Singkatnya, langkah-langkah ini adalah (1) mengembangkan dan mempertahankan SCM dari suatu sistem yang, minimal, mencakup semua “penyebab umum” yang diamati dan tidak diamati antara paparan dan hasil; (2) memblokir semua jalur non-kasual antara paparan dan hasil melalui penyesuaian statistik; dan (3) menyimpulkan efek kausal dari paparan.
pada hasil
, jika paparannya bersifat eksogen dan monotonik.
Karibu boreal di Kanada
Populasi rusa kutub hutan boreal (selanjutnya disebut “rusa kutub boreal” atau “karibu”) tersebar di sebagian besar hutan boreal Kanada dan terdaftar dalam Undang-Undang Spesies Berisiko Kanada sebagai Terancam (Environment and Climate Change Canada, 2020 ). Rusa kutub sangat penting bagi budaya dan identitas masyarakat Suku Bangsa Pertama Kanada, dan hak berburu mereka dilindungi berdasarkan perjanjian dan konstitusi Kanada.
Rusa kutub boreal sebagian besar hidup di hutan, hidup dalam kelompok kecil, dan tidak melakukan migrasi musiman jarak jauh seperti rusa kutub di tanah tandus ( Rangifer tarandus groenlandicus ). Mereka umumnya menempati hutan cemara ( Picea spp.) atau pinus ( Pinus spp.) dengan produktivitas rendah , yang menyediakan tempat berlindung sebagian dari predator (terutama serigala [ Canis lupus ]; Environment Canada, 2011 ). “Persaingan nyata yang dimediasi oleh gangguan” secara umum dianggap sebagai ancaman paling signifikan terhadap persistensi karibu (Neufeld et al., 2021 ), di mana perubahan habitat meningkatkan vegetasi seral awal yang lebih disukai oleh ungulata lain seperti rusa besar ( Alces alces ) dan rusa ( Odocoileus spp.), dan pada gilirannya, populasi predator meningkat dan secara tidak proporsional memengaruhi karibu karena produktivitas yang relatif rendah dari yang terakhir (misalnya, Mumma et al., 2018 ; Serrouya et al., 2015 ; Superbie et al., 2022 ; Wittmer et al., 2013 ). Efek ini kemungkinan lebih jelas di mana produktivitas lanskap lebih tinggi karena vegetasi seral awal diharapkan merespons gangguan dengan lebih kuat (Neufeld et al., 2021 ; Serrouya et al., 2021 ).
Caribou boreal di banyak bagian Kanada telah menurun selama beberapa dekade (Environment Canada, 2011 ), dan strategi pemulihan pertama kali diterbitkan pada tahun 2012 dan kemudian diubah pada tahun 2020 (Environment Canada, 2012 ; Environment and Climate Change Canada, 2020 ). Selain mengidentifikasi atribut biofisik habitat kritis, strategi pemulihan juga memperkenalkan pengelolaan habitat yang “terganggu” sebagai persyaratan habitat kritis. Habitat terganggu didefinisikan sebagai fitur antropogenik yang dapat diidentifikasi pada citra Landsat 1:50.000, dengan jarak 500 m, serta area yang terbakar dalam 40 tahun terakhir (Environment and Climate Change Canada, 2020 ). Untuk strategi pemulihan, hubungan yang diamati antara ukuran habitat yang terganggu dan perekrutan karibu (yaitu, rasio sub-dewasa terhadap betina dewasa yang “direkrut” ke dalam populasi pengembangbiakan) di antara sampel rentang karibu boreal digunakan untuk menetapkan persyaratan habitat kritis <35% habitat yang terganggu dalam setiap rentang karibu, metrik yang dilampaui di antara sebagian besar rentang Kanada. Seperti disebutkan di atas dan oleh yang lain (Sleep & Loehle, 2010 ), mengelola hubungan observasional tersebut berisiko mencampurkan kekuatan prediktifnya dengan atribusi kausal. Data demografi dan habitat karibu Untuk studi ini, saya menggunakan data demografi karibu, gangguan, dan batas wilayah studi dari Johnson et al. ( 2020 ), memperkirakan rekrutmen dan batas wilayah studi dari angka-angka yang tidak tersedia di tempat lain. Data ini mencakup 58 wilayah studi (Gambar 2 ) di seluruh wilayah jelajah karibu boreal Kanada, dengan data dikumpulkan antara tahun 1997 dan 2017. Analisis dibobot berdasarkan jumlah tahun observasi rekrutmen untuk setiap wilayah studi (total 159 observasi). Hanya rekrutmen rata-rata yang tersedia untuk wilayah studi dengan observasi >1 tahun (Johnson et al., 2020 ).
Saya menggunakan data gangguan habitat terkini hingga 2015 yang disusun oleh Environment and Climate Change Canada (diakses di: https://open.canada.ca/data/dataset/a71ab99c-6756-4e56-9d2e-2a63246a5e94 ) menggunakan metode yang dijelaskan oleh Pasher et al. ( 2013 ). Saya menggunakan metrik gangguan agregat (yaitu, fitur antropogenik yang dibatasi oleh 500 m, serta area terbakar <40 tahun), tetapi juga menghitung secara terpisah kepadatan fitur linear (misalnya, jalan, garis seismik) di setiap area studi, serta persentase setiap rentang yang ditutupi oleh blok tebangan kehutanan. Sesuai Pasher et al. ( 2013 ), blok tebangan dimasukkan dalam analisis jika terlihat pada citra Landsat 1:50.000. Karena gangguan sering kali saling tumpang tindih, saya tetapkan jenis gangguan dalam prioritas berikut: fitur linier, jika tidak fitur antropogenik poligonal, jika tidak kebakaran terkini.
Saya mengkarakterisasi produktivitas lanskap menggunakan MODIS 500-m Enhanced Vegetation Index (EVI) yang diakses melalui Google Earth Engine ( https://earthengine.google.com ) dan Google Colab ( https://colab.research.google.com ), menggunakan skrip Python untuk mengekstrak koleksi gambar untuk Kanada pada bulan Juli setiap tahun dari 2011 hingga 2015 pada resolusi 1 km (Gambar 2 ). Gambar raster ini kemudian dirata-ratakan menggunakan QGIS 3.2 ( QGIS.org , 2023 ) untuk mengkarakterisasi produktivitas lanskap. Lima tahun dirata-ratakan untuk mengatasi variasi tahunan dan tutupan awan.
Menilai atribusi kausal gangguan habitat dalam sistem karibu
Saya mengajukan pertanyaan kontra-faktual berikut: apakah penurunan yang diamati dalam kelimpahan rusa kutub (seperti yang diperkirakan dari perekrutan remaja) di antara subpopulasi di area studi dengan gangguan habitat tinggi akan terjadi seandainya area studi tersebut memiliki gangguan habitat rendah dan, dengan demikian, apakah subpopulasi yang meningkat secara stabil yang diamati di area studi dengan gangguan habitat rendah akan tetap demikian seandainya mereka mengalami gangguan habitat yang tinggi.
Saya menggunakan prosedur yang dijelaskan dalam Boreal caribou di Kanada untuk memperkirakan: (1) atribusi kausal dari ukuran agregat gangguan habitat, dengan asumsi efek langsung dan kausal pada rekrutmen karibu, seperti yang disajikan dalam Environment Canada ( 2011 ) tetapi menggunakan data terbaru yang disusun oleh Johnson et al. ( 2020 ) dan (2) model alternatif yang memisahkan gangguan habitat dan produktivitas lanskap untuk menilai jalur fungsional berbeda yang memengaruhi sistem karibu.
Model gangguan agregat
Mengikuti Environment Canada ( 2011 , 2012 ), saya menggunakan tingkat rekrutmen 29 anak sapi:100 sapi sebagai ambang biner untuk mengklasifikasikan subpopulasi sebagai populasi yang stabil dan meningkat
) atau menurun
; Environment Canada, 2008 ), dan ambang batas kebijakan sebesar 35% untuk mengklasifikasikan gangguan habitat sebagai gangguan rendah (
) atau tinggi (
). Saya juga melakukan analisis sensitivitas untuk menentukan dampak kebijakan gangguan habitat agregat yang berbeda dengan memvariasikan ambang gangguan ±30% dengan kelipatan 5%.
Model kausal tersirat dari hubungan agregat gangguan-rekrutmen diwakili oleh dua simpul ( gangguan paparan dan rekrutmen hasil ) dan satu busur terarah (Gambar 3 ). Bukti observasional yang menghubungkan rekrutmen dengan gangguan habitat dalam konteks model yang diasumsikan ini dapat dianggap kausal karena paparan bersifat eksogen, dan saya berasumsi monotonisitas (yaitu, gangguan tinggi tidak menguntungkan rekrutmen karibu di area studi mana pun).

Saya membuat parameter SCM sebagai model jaringan Bayesian deterministik, yang ditulis menggunakan paket R (versi 4.4.2; Tim Inti R, 2024 ) bnlearn (versi 5.0.1; Scutari dan Ness, 2020 ) menggunakan data berkode biner (Gambar 4 ). Jaringan Bayesian mengkodekan dependensi bersyarat di antara variabel terdiskritisasi, seperti yang direpresentasikan oleh simpul yang dihubungkan oleh busur terarah dalam DAG (Pearl, 1988 ). Dalam contoh ini, hasil biner (rekrutmen tinggi versus rendah) sepenuhnya ditentukan oleh frekuensi relatif gangguan habitat tinggi atau rendah. Saya menetapkan bukti dalam model jaringan pada gangguan tinggi (
) dan mengamati kemungkinan yang berhubungan dengan rendahnya rekrutmen (
) untuk menghitung probabilitas faktual
(Persamaan 3 ). Di sini,
setara dengan proporsi pengamatan di kuadran kanan bawah Gambar 4 dalam kaitannya dengan jumlah total pengamatan di kuadran kanan bawah dan atas.
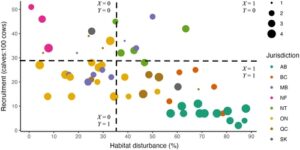
Untuk menghitung probabilitas kontrafaktual
(Persamaan 4 ), Saya menetapkan bukti dalam model jaringan pada gangguan rendah (
) dan kembali mengamati kemungkinan yang berhubungan dengan rendahnya rekrutmen (
). Di Sini,
setara dengan proporsi pengamatan di kuadran kiri bawah Gambar 4 dalam kaitannya dengan jumlah total pengamatan di kuadran kiri bawah dan atas.
Dengan probabilitas faktual dan kontra-faktual ini, saya kemudian menggunakan Persamaan ( 8-10 ) untuk menghitung probabilitas kausalitas perlu, cukup, dan perlu dan cukup.
Model gangguan terurai dan produktivitas lanskap
Saya mengembangkan hipotesis kausal alternatif untuk menjelaskan tingkat perekrutan karibu berdasarkan sumber gangguan habitat yang berbeda (yaitu, fitur linier, blok pemotongan, dan api) dan produktivitas lanskap.
Saya memilih ambang batas untuk membedakan gangguan tinggi dari rendah untuk model terurai guna membedakan hubungan univariat dengan lebih baik. Tidak ada pengamatan rekrutmen tinggi (yaitu, ≥29 anak sapi:100 sapi) di area studi dengan kepadatan >0,2 km/km 2 fitur linier, jadi saya menggunakan ambang batas tersebut untuk membedakan kepadatan fitur linier tinggi dari rendah. Hal yang sama berlaku ketika >10% area studi diklasifikasikan sebagai blok tebang hutan. Untuk kebakaran baru-baru ini, semua kecuali satu area studi dengan rekrutmen tinggi dikaitkan dengan kondisi di mana <40% area telah terbakar dalam 40 tahun (Dehcho North, NT). Untuk produktivitas primer, hanya satu area studi dengan rekrutmen tinggi dikaitkan dengan EVI rata-rata > 3500 (Bloodvein, MB). Nilai-nilai ini memaksimalkan
untuk faktor-faktor masing-masing dan oleh karena itu atribusi kausal.
Saya berhipotesiskan struktur kausal sederhana dengan keempat faktor sebagai induk independen dari rekrutmen dan menyesuaikan data ke jaringan. Penyertaan api sebagai faktor menghasilkan sel kosong dalam tabel probabilitas kondisional (terlepas dari ambang batas), dan faktor ini memiliki informasi timbal balik kondisional terendah dengan simpul rekrutmen target, jadi saya menghapus busur kausal (Gambar 5 ). Jaringan kausal ini memenuhi kriteria eksogenitas (yaitu, tidak ada busur masuk ke paparan) dan saya mengasumsikan monotonisitas dalam efek faktor. Saya kemudian menggunakan Persamaan ( 3 , 4 , 8-10 ) untuk menghitung probabilitas kausalitas yang diperlukan, cukup, dan perlu dan cukup.
HASIL
Model gangguan agregat
Berdasarkan model gangguan agregat, probabilitas bahwa gangguan habitat yang tinggi (>35%) diperlukan (PN) untuk menyebabkan rekrutmen rendah adalah 20,8% (Tabel 2 ), lebih rendah daripada probabilitas bahwa itu cukup (PS = 53,7%). Secara kolektif, probabilitas bahwa gangguan habitat yang tinggi diperlukan dan cukup (PNS) untuk menyebabkan rekrutmen rendah adalah 17,6%.
| Model | Pengkondisian | Bukti | Probabilitas faktual suatu peristiwa ( p 1 ) | Probabilitas kontrafaktual suatu peristiwa ( p 0 ) | Probabilitas keharusan (PN %) | Probabilitas kecukupan (PS %) | Probabilitas kebutuhan dan kecukupan (PNS %) |
|---|---|---|---|---|---|---|---|
| Gangguan agregat | Tidak ada | Perawatan <65% tidak terganggu | 0.848 | 0.672 | 20.8 | 53.7 | 17.6 |
| Produktivitas primer | Tidak ada | >3500 | 0.980 | 0,677 tahun | 31.0 | 93.9 | 30.3 |
| Gangguan terpisah | Tidak ada | Linier >0,02 | 1.000 | 0.742 | 25.8 | 100.0 | 25.8 |
| Blok potong >10% | 1.000 | 0.803 | 19.7 | 100.0 | 19.7 | ||
| Blok potong >10%, Linier >0,02 | 1.000 | 0.705 | 29.5 | 100.0 | 29.5 | ||
| EVI Tinggi | Linier >0,02 | 1.000 | 0,970 | 3.0 | 100.0 | 3.0 | |
| Blok potong >10% | 1.000 | 0,977 tahun | 2.3 | 100.0 | 2.3 | ||
| Blok potong >10%, Linier >0,02 | 1.000 | 0,966 tahun | 3.4 | 100.0 | 3.4 | ||
| EVI rendah | Linier >0,02 | 1.000 | 0,515 | 48.5 | 100.0 | 48.5 | |
| Blok potong >10% | 1.000 | 0.631 | 36.9 | 100.0 | 36.9 | ||
| Blok potong >10%, Linier >0,02 | 1.000 | 0.446 | 55.4 | 100.0 | 55.4 |
Singkatan: EVI, Enhanced Vegetation Index.
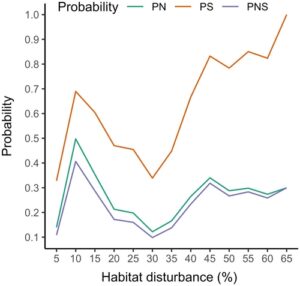
Probabilitas kausalitas sensitif terhadap ambang batas gangguan habitat, dengan minimum terjadi pada 30% (Gambar 6 ). Pada ambang batas gangguan yang meningkat, PS meningkat menjadi 1 karena jumlah pengamatan rentang gangguan tinggi dengan rekrutmen tinggi menjadi 0 di atas ambang batas 60%. Hasil pada ambang batas 5%–15% lebih bervariasi, dengan ukuran sampel kecil dan rasio pergeseran rentang dengan rekrutmen tinggi versus rendah.
Model gangguan dan produktivitas yang terdisagregasi
Kepadatan fitur linear yang tinggi (yaitu, >0,2 km/km 2 ) memiliki PN sebesar 25,8% (Tabel 1 ) dan PS = 100% (mengingat tidak ada pengamatan subpopulasi yang meningkat secara stabil dalam rentang dengan kepadatan fitur linear yang tinggi). PNS sama dengan PN (25,8%) karena PS = 100%. PN dan PNS dari blok potong >10% dari suatu rentang adalah 19,7%. Efek kumulatif dari kedua sumber gangguan memiliki PN dan PNS sebesar 29,5%.
Produktivitas primer, yang diukur dengan rata-rata EVI Juli, memiliki PNS sebesar 30,3%, melebihi kepadatan fitur linier tinggi atau blok tebangan >10% dari suatu rentang. Stratifikasi berdasarkan produktivitas tinggi versus rendah mengubah kemungkinan sebab akibat. Efek kumulatif fitur linier dan blok tebangan (gangguan habitat tinggi) dikaitkan dengan PNS rendah (<3,5%) di area produktivitas tinggi. Namun di area produktivitas rendah, gangguan habitat tinggi dikaitkan dengan PNS tinggi (55,4%).
DISKUSI
Analisis kebijakan lingkungan memerlukan pengetahuan tentang hubungan sebab-akibat, tetapi bukti eksperimental seringkali sulit diperoleh (Cucurachi dan Suh, 2017 ). Dalam keadaan ini, adalah umum untuk melakukan analisis korelatif dari data observasional dan kemudian berspekulasi tentang hubungan kausal. Tetapi mendasarkan intervensi manajemen pada spekulasi kausal bisa mahal, terutama jika intervensi memiliki biaya langsung atau peluang yang tinggi dan hasil yang diinginkan tidak terwujud karena kelemahan mendasar dalam logika kausal. Pertanyaan utama dalam penalaran retrospektif tentang kausalitas dalam sistem ekologi adalah apakah hasil buruk yang diamati akan berbeda jika paparan tidak terjadi (Coetzee dan Gaston, 2021 ). Estimasi semacam itu memberikan dasar yang lebih kuat untuk merancang intervensi kebijakan untuk membalikkan hasil yang merugikan. Seperti yang ditunjukkan di sini, kemajuan dalam analisis kausal menyediakan alat analitis untuk memperkirakan secara retrospektif kontribusi kausal dari berbagai faktor tanpa adanya data eksperimen.
Tentu saja, tidak ada analisis data observasional yang dapat menandingi kekuatan uji coba kontrol acak untuk menetapkan kausalitas (Cartwright, 2010 ), dan model kausal yang hanya berdasarkan observasi tetap, pada intinya, analisis korelasi, yang menerapkan kehati-hatian interpretasi yang terkenal (Cliff, 1983 ). Beralih dari model prediktif ke model kausal, sebagian, memerlukan peralihan dari kehati-hatian ini ke asumsi eksplisit (misalnya, kehati-hatian akan adanya unsur pengganggu dalam interpretasi model korelasional menjadi asumsi eksogenitas dalam model kausal). Akibatnya, penerimaan inferensi yang diperoleh dari model kausal bergantung pada transparansi dan kewajaran asumsi mereka seperti pada diagnostik statistik mereka. Saya menyarankan bahwa mempertimbangkan model sebagai hipotesis kausal dan mengevaluasi konsekuensinya di bawah asumsi yang ketat merupakan peningkatan daripada secara implisit mengasumsikan kausalitas dalam hubungan statistik. Lebih lanjut, saya menyarankan bahwa PN, PS, dan PNS adalah estimasi sederhana dan intuitif mengenai implikasi kontrafaktual dari hipotesis tersebut dan memungkinkan peneliti untuk mengeksplorasi pertanyaan retrospektif tentang atribusi kausal dengan bukti observasional yang tersedia.
Dalam studi ini, pertanyaan kontrafaktual yang diajukan adalah apakah penurunan yang diamati dalam kelimpahan karibu di antara subpopulasi di area studi dengan gangguan habitat tinggi akan terjadi jika area studi tersebut malah memiliki gangguan habitat rendah dan, dengan demikian, apakah subpopulasi yang meningkat secara stabil yang diamati di area studi dengan gangguan habitat rendah akan tetap demikian jika mereka terpapar pada gangguan habitat yang tinggi. Sementara yang lain telah mengekstrapolasi model berbasis regresi untuk menyimpulkan secara retrospektif atau prospektif respons populasi karibu di bawah rezim manajemen alternatif (misalnya, Johnson et al., 2020 ; Stewart et al., 2020 ; Serrouya et al., 2011 ), tidak ada yang mengadopsi pendekatan kausal-kontrafaktual eksplisit seperti yang saya lakukan di sini.
Dengan model kausal tersirat dan ambang batas kebijakan gangguan habitat saat ini sebesar 35%, gangguan habitat agregat ditemukan memiliki atribusi kausal yang rendah (PNS = 17,6%), yang menunjukkan bahwa tujuan pemulihan saat ini untuk mengurangi gangguan agregat menjadi <35% rata-rata memiliki probabilitas rendah untuk berhasil meningkatkan rekrutmen ke tingkat yang konsisten dengan populasi yang meningkat secara stabil. Alasan untuk PNS yang rendah ini adalah bahwa, sementara banyak subpopulasi di area studi gangguan habitat tinggi memang menurun, sebagian besar subpopulasi dalam kondisi gangguan rendah juga menurun. Selain itu, ada juga subpopulasi yang meningkat secara stabil di area studi dengan gangguan habitat tinggi. Pengamatan keadaan ini bertentangan dengan hubungan gangguan habitat-rekrutmen yang diprediksi, dan frekuensi relatifnya menginformasikan interpretasi kausal model. Dalam kasus ini, model memiliki PN yang relatif rendah tetapi PS yang lebih tinggi. Ini menunjukkan bahwa gangguan habitat jauh dari penting (PN = 20,8%) tetapi dapat memadai (PS = 53,7%) untuk menyebabkan rekrutmen karibu yang rendah dengan sendirinya. Hal ini juga menunjukkan adanya beberapa jalur kausal, yang mana gangguan agregat hanya merupakan salah satunya. Probabilitas kausalitas dihitung dari input biner dan karenanya tunduk pada efek ambang batas. Dalam studi ini, persentase gangguan habitat baik di bawah maupun di atas ambang batas kebijakan 35% dikaitkan dengan probabilitas kausalitas yang lebih tinggi. Dalam praktiknya, probabilitas kausalitas minimal ketika Dan paling mirip karena bukti hubungan kausal pasti lemah ketika suatu hasil terjadi secara kontrafaktual pada frekuensi yang sama dengan yang terjadi secara faktual. Ketika probabilitas hasil kontrafaktual menurun relatif terhadap hasil faktual, probabilitas kausalitas meningkat. Metode untuk memperluas probabilitas kausalitas ke variabel kontinu mulai bermunculan (Kawakami et al., 2024 ). Merevisi model kausal untuk memisahkan sumber gangguan habitat ke dalam jalur yang berbeda menuju penurunan menunjukkan bahwa ada kondisi gangguan yang, berdasarkan data yang tersedia saat ini, tidak sesuai dengan populasi rusa kutub yang terus meningkat; khususnya, kepadatan fitur linear >0,2 km/km 2 dan/atau area di mana blok penebangan hutan mencakup >10% dari area jangkauan. Kedua ambang batas dikaitkan dengan PS = 100%, yang menunjukkan tidak ada bukti bahwa subpopulasi rusa kutub dapat stabil dalam kondisi yang melampaui ambang batas ini.
Fitur linear dan blok pemotongan hutan umumnya dikaitkan dengan jalur yang berbeda menuju penurunan dalam sistem karibu, dengan blok pemotongan yang menghasilkan vegetasi seral awal diasumsikan mendorong persaingan yang jelas dan fitur linear menyediakan koridor perjalanan yang meningkatkan permeabilitas habitat yang sebaliknya berfungsi sebagai tempat perlindungan predator untuk karibu (misalnya, DeMars & Boutin, 2018 ; Dickie et al., 2017 ; Mumma et al., 2018 ). Kedua faktor ini secara luas telah dikarakterisasi sebagai respons “numerik” dan “fungsional” dari sistem predator-mangsa terhadap perubahan habitat yang disebabkan oleh manusia, dan analisis ini menunjukkan bahwa kepadatan fitur linear memiliki atribusi kausal yang lebih kuat (PNS 25,8% vs. 19,7%), dengan efek kumulatif dari melampaui kedua ambang batas menjadi lebih tinggi (PNS = 29,7%). Ini adalah studi pertama yang menetapkan atribusi kausal independen pada faktor-faktor ini tetapi konsisten dengan kesimpulan Mumma et al. ( 2018 ), yang menemukan, di British Columbia timur laut, lebih banyak dukungan untuk efek langsung fitur linear pada tumpang tindih karibu-serigala daripada kompetisi nyata numerik atau spasial.
Proporsi area studi yang baru-baru ini terbakar oleh kebakaran hutan bukan merupakan faktor signifikan dalam analisis. Studi terbaru menunjukkan bahwa kebakaran mungkin bukan pendorong yang sepenting dalam penurunan populasi rusa kutub boreal seperti yang diperkirakan sebelumnya (DeMars et al., 2019 ; Johnson et al., 2020 ; Neufeld et al., 2021 ). Namun, hal ini mungkin tidak berlaku jika perubahan iklim meningkatkan frekuensi perilaku kebakaran ekstrem di Kanada seperti yang diprediksi (Wang et al., 2015 ).
Mirip dengan Serrouya et al. ( 2021 ) dan Neufeld et al. ( 2021 ), saya menemukan bahwa produktivitas primer merupakan faktor signifikan dalam sistem karibu. Probabilitas kausalitas yang diperlukan dan cukup dari produktivitas primer (30,3%) serupa dengan gabungan sumber antropogenik dari fitur linear dan blok tebangan (29,5%). Atribusi kausal lebih tinggi untuk sumber gangguan antropogenik di area dengan produktivitas primer rendah, yang menunjukkan bahwa faktor selain gangguan habitat mungkin penting di habitat yang lebih produktif. Ini dapat mencakup perluasan rusa berekor putih ( Odocoileus virginianus ) yang terlepas dari perubahan habitat yang signifikan (misalnya, melalui pemanasan iklim; Dawe & Boutin, 2016 ), yang juga berkontribusi pada persaingan yang tampak.
Gangguan habitat sering disebut sebagai penyebab “utama” penurunan populasi rusa kutub karena efek berjenjang pada vegetasi dan perubahan predator-mangsa yang ditimbulkannya (misalnya, Baillie-David et al., 2024 ; DeMars et al., 2023 ; Festa-Bianchet et al., 2011 ; Nagy-Reis et al., 2021 ), yang menyebabkan beberapa pihak menyimpulkan bahwa pemulihan habitat akan cukup untuk meniadakan perlunya tindakan berbasis populasi yang intensif seperti pengendalian predator (Environment and Climate Change Canada, 2020 ; Lamb et al., 2024 ). Namun, atribusi kausal (PNS) dari faktor-faktor terkait gangguan yang saya pertimbangkan hanya 29,5% di seluruh rentang, yang menunjukkan bahwa kesimpulan seperti itu mungkin prematur dan bahwa ada faktor sistematis yang tidak teramati (menunjukkan model kausal yang berbeda) atau faktor stokastik yang beroperasi dalam sistem karibu boreal yang dapat membatasi efektivitas pemulihan habitat atau regenerasi alami dalam membangun kembali populasi karibu yang mandiri. Faktor-faktor ini dapat mencakup variasi spasial atau temporal atau pergeseran pasokan hijauan, penyakit/parasit, hasil buruan pemburu, perpindahan, atau lainnya (DeMars et al., 2023 ). Pemulihan karibu yang berhasil akan memerlukan pemahaman dan pengelolaan yang lebih baik dari faktor-faktor lain yang menyebabkan karibu menurun.