
Abstrak
Morfologi makanan sangat beragam pada Carnivoraformes (Karnivora hidup dan kerabat batangnya) selama Kenozoikum (66 juta tahun terakhir) saat mereka berevolusi untuk menangkap, menangani, dan memproses makanan hewani dan nabati baru. Kami menggunakan morfometrik geometri 3D, keuntungan mekanis, dan analisis elemen hingga untuk menguji hubungan evolusi antara bentuk mandibula dan fungsi biomekanik saat subklade secara independen melakukan transisi dari makanan mesokarnivora (50%–70% bahan hewani) menjadi hiperkarnivora (>70% bahan hewani) dan makanan osteofag (pemrosesan tulang yang substansial). Kami menemukan bahwa bentuk mandibula berkorelasi dengan kategori makanan ini, dengan estimasi keuntungan mekanis, dan dengan stres dan regangan yang disebabkan oleh interaksi antara beban gigi taring dan posisi temporalis relatif terhadap carnassial. Pemisahan kategori makanan kemungkinan terkait dengan perbedaan bentuk mandibula terkait bentuk kondilus, bentuk perlekatan otot, panjang carnassial, dan panjang serta kelengkungan ramus horizontal. Hal ini pada gilirannya terkait dengan estimasi keuntungan mekanis karena yang paling erat kaitannya adalah yang terkait dengan pemanjangan lengan pengungkit temporalis dan pemendekan mandibula dan lengan pengungkit titik gigitan. Perbedaan tekanan dan regangan kemungkinan terkait dengan variasi di bagian distal (atau rostral) mandibula yang terkait dengan mangsa dengan ukuran yang berbeda (mesokarnivora biasanya memangsa yang lebih kecil dari ukuran tubuhnya sendiri, sedangkan hiperkarnivora memangsa yang sama atau lebih besar dari dirinya sendiri). Taksa mesokarnivora, secara rata-rata, memiliki tekanan dan regangan yang lebih tinggi pada mandibula daripada kelompok makanan lainnya.
1. PENDAHULUAN
Saat ini, klade Carnivoraformes, kelompok karnivora mamalia yang masih hidup yang mencakup kucing, anjing, beruang, rakun, dan taksa lain serta kerabat batangnya yang telah punah, menunjukkan berbagai macam ekologi makanan, di antaranya adalah mesokarnivora (makanan yang terdiri dari 50%–70% bahan hewani), hipokarnivora (<50% bahan hewani), hiperkarnivora (>70% bahan hewani), dan hiperkarnivora pemecah tulang (Radinsky, 1982 ; Roemer et al., 2009 ; Werdelin & Gittleman, 1996 ). Keragaman makanan mereka disertai dengan variasi luas dalam morfologi mandibula mereka. Mesokarnivora, seperti rubah merah, Vulpes vulpes , dan genet umum, Genetta genetta , sering memiliki rahang yang panjang dan ramping untuk menangkap mangsa kecil dengan cepat dan mempertahankan permukaan penghancur di bagian kaudal bilah carnassial (Friscia et al., 2007 ; Roemer et al., 2009 ; Van Valkenburgh, 2007 ). Hipokarnivora, seperti beruang hitam, Ursus americanus , dan luak Eropa, Meles meles , memiliki rahang dengan tempat perlekatan otot yang besar baik pada fossa masseterik maupun prosesus koronoid, permukaan geser kecil pada carnassial mereka, dan area penghancuran molar yang lebih besar di belakang permukaan geser carnassial (Greaves, 1983 ; Meloro & Raia, 2010 ; Van Valkenburgh, 2007 ). Hiperkarnivora, seperti harimau, Panthera tigris , dan musang ekor panjang, Mustela frenata , memiliki rahang pendek untuk mengurangi kekuatan gigitan, prosesus koronoid yang lebih panjang dan kondilus, gigi taring, simfisis mandibula, dan permukaan geser carnassial yang lebih besar (Van Valkenburgh, 1991 , 1999 , 2007 ; Van Valkenburgh et al., 2004 ). Hiperkarnivora pemecah tulang, seperti hyena tutul, Crocuta crocuta , memiliki gigi yang lebih kuat dan perpindahan dorsoventral ramus asenden dan ramus horizontal mandibula (Ferretti, 2007 ; Tseng, 2013 ; Tseng et al., 2011 ; Tseng & Wang, 2010 ; Van Valkenburgh, 1999 , 2007 ). Morfologi ini bervariasi dalam kinerja fungsionalnya sebagaimana diukur dengan metrik independen seperti keuntungan mekanis atau total tegangan dan regangan.
Carnivoraformes terdiri dari beberapa subklade yang masih ada dan sejumlah taksa punah yang berkerabat dekat dengan mereka (Gambar 1 ; Baskin, 1998 ; Flynn et al., 2005 ; Spaulding & Flynn, 2012 ; Wozencraft, 1989 ). Kelompok parafiletik Miacidae, yang merupakan anggota kelompok paling awal dan berkerabat dekat dengan kelompok yang masih ada, memiliki berbagai spesialisasi mesokarnivora dan sebagian besar mempertahankan rencana gigi plesiomorfik Carnivoraformes (I3/3, C1/1, P4/4, M3/3). Genus Vulpavus mewakili miacids dalam penelitian ini (Radinsky, 1982 ; Solé et al., 2016 ; Sole & Smith, 2013 ; Spaulding & Flynn, 2012 ). Kisaran kecil dalam ukuran tubuh dan pola makan mesokarnivora yang ada di mana-mana pada karnivoraform awal telah dikaitkan dengan kendala yang diberlakukan oleh pengecualian kompetitif dari kelompok karnivora mamalia lain yang secara filogenetik independen dan sekarang sudah punah, termasuk Mesonychia, Hyaenodonta, dan Oxyaenodonta (Martin, 1989 ; Van Valkenburgh, 1988 , 1999 , 2007 ). Pada akhir Eosen, sekitar 42 juta tahun yang lalu, anggota kelompok mahkota Carnivora mulai bercabang menjadi apa yang akan menjadi Canidae, Mustelidae, kelompok awal Feliformia, dan kelompok yang punah seperti Amphicyonidae. Pada zaman Miosen 20 juta tahun yang lalu, semua keluarga Karnivora yang masih ada telah berevolusi, termasuk Felidae, Hyaenidae, Herpestidae, Ursidae, Procyonidae, dan Viverridae (Gregory & Hellman, 1939 ; Hunt Jr, 1998 ; Martin, 1989 ; Van Valkenburgh, 1991 , 1999 ; Wozencraft, 1989 ).
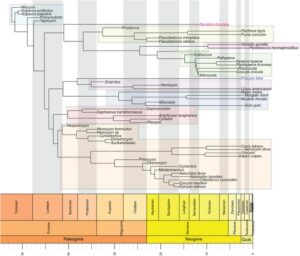
Mesokarnivora generalis merupakan ekologi makanan leluhur Carnivoraformes, dan spesialisasi makanan lainnya (hipokarnivora, hiperkarnivora, dan hiperkarnivora pemecah tulang) berevolusi seiring dengan diversifikasi klad (Greaves, 1985 ; Martin, 1989 ; Radinsky, 1982 ; Van Valkenburgh, 2007 ). Perbedaan tertentu dalam bentuk mandibula telah disimpulkan menjadi konsekuensi fungsional dari spesialisasi ini, dan jika demikian, kita akan menduga perubahan filogenetik dalam bentuk akan dikaitkan dengan perubahan dalam kinerja biomekanik (Davis, 2014 ; Dullemeijer & Barel, 1977 ; Greaves, 1985 ; Heinrich, 1996 ; Herring, 1993 ; Meloro et al., 2015 ; Radinsky, 1982 ; Werdelin & Gittleman, 1996 ). Karena spesialisasi hipokarnivora, hiperkarnivora, dan retakan tulang telah berevolusi secara independen beberapa kali dalam subklade karnivoraform yang berbeda, kami berharap bentuk mandibula akan menunjukkan konvergensi ekomorfologi (ekomorf yang signifikan secara statistik) (Christiansen & Adolfssen, 2005 ; Goswami & Polly, 2010 ; Meloro & Raia, 2010 ; Van Valkenburgh, 2007 ). Misalnya, beberapa penulis berpendapat bahwa ekomorf “felid” hiperkarnivora yang saat ini ditemukan tidak hanya pada kucing tetapi juga ditemukan pada kaniform yang telah punah seperti mustelid Megalictis dan canid Enhydrocyon yang morfologinya seperti kucing telah diusulkan sebagai hasil dari mereka mengisi peran ekologis seperti kucing selama “kesenjangan kucing” Amerika Utara (27–17 juta tahun lalu) (Christiansen, 2008 ; Goswami et al., 2011 ; Meloro et al., 2015 ; Van Valkenburgh, 1991 , 1999 , 2007 ). Contoh lain adalah ekomorf osteofagus “hyaenid” yang ditemukan di Hyaenidae dan beberapa Canidae (khususnya subfamili Borophaginae) yang terdiri dari tengkorak berbentuk kubah, ramus menaik yang bergeser dari ramus horizontal, prosesus koronoid yang besar dan sejajar secara vertikal, dan gigi yang lebih kuat (Ferretti, 2007 ; Tseng, 2013 ; Tseng et al., 2011 ; Van Valkenburgh, 2007 ). Jika ekomorf bentuk ini benar-benar dioptimalkan untuk persyaratan biomekanik dari strategi diet masing-masing, maka kami berharap untuk melihat perubahan berkorelasi dalam bentuk mandibula dan fungsi biomekanik karena subklade yang berbeda telah mengembangkan spesialisasi ini.
Dalam makalah ini kami menguji proposisi-proposisi ini menggunakan pemindaian digital tiga dimensi (3D) dari mandibula karnivoraform yang diambil sampelnya dari beberapa transisi independen dari pola makan mesokarnivora leluhur ke pola makan yang lebih berkembang untuk menguji hubungan antara pola makan, bentuk mandibula, dan kinerja fungsional. Kami berhipotesis bahwa filogeni akan memiliki korelasi yang kuat dengan variasi bentuk mandibula, tetapi pola makan dan kinerja fungsional yang dikaitkan dengan ekomorf akan memiliki korelasi yang lebih kuat yang melampaui aspek bentuk yang spesifik secara filogenetik. Kami akan menggunakan analisis elemen hingga, pengukuran keuntungan mekanis, dan analisis morfometrik geometris untuk mendokumentasikan pola gaya gigitan dan evolusi stres dan regangan mandibula pada mandibula karnivoraform. Pola-pola ini akan dianalisis dalam konteks filogenetik untuk menentukan apakah transisi antara mesokarnivora dan hiperkarnivora secara universal dikaitkan dengan pergeseran dari bentuk mandibula stres tinggi ke bentuk stres yang lebih rendah. Taksa hipokarnivora juga disertakan dalam penelitian ini untuk memberikan kontras dengan taksa hiperkarnivora dan pemecah tulang.
2 BAHAN DAN METODE
2.1 Singkatan institusi dan terminologi kedokteran gigi
Singkatan institusional: Museum Sejarah Alam Amerika (AMNH), Koleksi Zooarchaeology William R. Adams Universitas Indiana (WRAZL), Museum Zoologi Universitas Michigan (UMMZ), Museum Sejarah Alam Florida Universitas Florida (UF), Museum national d’Histoire naturelle (MNHN), Museum Sejarah Alam Hefner (MUZO), Museum Zoologi Vertebrata Universitas California Berkeley (MVZ), Museum Sejarah Alam Idaho (IMNH), Universitas Negeri Carolina Utara (NCSU), Musem van Naturlijke Historie, Leiden (MNHL), Museum Sejarah Alam Nasional Smithsonian Institution (USNM), Museum Universitas Arkansas (UAM), Museum Sejarah Alam San Diego (SDMNH), dan Museum Yale Peabody (YPM).
Istilah dan singkatan kedokteran gigi pada umumnya mengikuti Bown dan Krause ( 1979 ). Di sini singkatan jenis gigi berikut digunakan, dengan huruf kecil menunjukkan gigi mandibula dan huruf besar menunjukkan gigi maksila: i, gigi seri; c, gigi taring; p, gigi premolar; m, gigi molar.
2.2 Spesimen
Sebanyak 54 spesimen dipilih untuk mengambil sampel sebanyak mungkin transisi filogenetik dari mesokarnivora ke salah satu ekologi diet khusus, yang mana 17 di antaranya masih ada dan sisanya telah punah. Setiap spesies diwakili oleh satu rahang dewasa sebagaimana ditentukan dengan memiliki gigi permanen penuh. Mandibula fosil sering kali tidak memiliki kondilus, prosesus koronoideus, atau gigi utama, dan dalam beberapa kasus, pengambilan sampel taksa fosil kami dibatasi oleh ketersediaan mandibula lengkap atau hampir lengkap yang tidak mengalami deformasi tafonomis. Rincian taksonomi dapat dilihat pada Tabel 1 . Pilihan kami mencakup 80% dari 15 famili Carnivoraformes yang masih ada dan punah, dengan hanya Ailuridae (panda merah, yang dominan omnivora), Eupleridae (radiasi eksklusif Malagasi), dan Prionodontidae (linsang) yang hilang (Martin, 1989 ; Van Valkenburgh, 2007 ).
| Taksa | Kelompok kladistik yang lebih besar | Keluarga | Ekologi | Koleksi | Nomor katalog. | Periode | Tipe data | Sumber |
|---|---|---|---|---|---|---|---|---|
| Adkrokuta | Karnivora | Suku Hyaenidae | Pemecah tulang | AMNH | 22.880 | Miosen | Fotogrametri | Kunjungan database dan koleksi |
| Aelurodon ferox | Karnivora | Anjing | Pemecah tulang | Angkatan Laut AS | 352.364 orang | Miosen | Fotogrametri | Kunjungan koleksi |
| Aelurodon taksoidea | Karnivora | Anjing | Pemecah tulang | AMNH | 67.008 orang | Miosen | Fotogrametri | Kunjungan koleksi |
| Aelurodon taksoidea | Kaniformia | Anjing | Pemecah tulang | YPM | VPPU 10635 | Miosen | Foto | Kunjungan koleksi |
| Aenocyon dirus | Karnivora | Anjing | Hiperkarnivora | YPM | Nomor Uraian 017990B | Zaman Pleistosen | Permukaan dipindai | Dibagikan |
| Amfibi longiramus | Karnivora | Amphicyonidae | Pemecah tulang | Bahasa Indonesia: UF | Wakil Presiden 019909 | Miosen | Permukaan dipindai dan difoto | Kunjungan koleksi |
| Anjing latrans | Karnivora | Anjing | Mesokarnivora | Universitas Amrik | 88–50-290 | Holosen | Dipindai CT | Sumber Morfo |
| Crocuta crocuta | Karnivora | Suku Hyaenidae | Pemecah tulang | MVZ | 165.169 tahun | Holosen | Permukaan dipindai | Sumber Morfo |
| Burung Cynarctus | Karnivora | Anjing | Hipokarnivora | Angkatan Laut AS | 3.524.050 | Miosen | Fotogrametri | Kunjungan koleksi |
| Bahasa Inggris Cynelos | Karnivora | Amphicyonidae | Mesokarnivora | Bahasa Indonesia: UF | Wakil Presiden 211624 | Miosen | Permukaan dipindai dan difoto | Kunjungan koleksi |
| Sinodesmus | Karnivora | Anjing | Mesokarnivora | YPM | VPPU 010412 | Oligosen | Permukaan dipindai | Dibagikan |
| Daphoenodon sp | Karnivora | Amphicyonidae | Pemecah tulang | Bahasa Indonesia: UF | Wakil Presiden 270828 | Miosen | Permukaan dipindai dan difoto | Kunjungan Koleksi |
| Daphoenus hartshornianus | Karnivora | Amphicyonidae | Mesokarnivora | Bahasa Indonesia: UF | Wakil Presiden 207948 | Oligosen | Permukaan dipindai dan difoto | Kunjungan koleksi |
| Desmocyon | Karnivora | Anjing | Mesokarnivora | AMNH | Nomor Telepon 49177 | Oligosen | Fotogrametri | Kunjungan koleksi dan database |
| Enhidrokion | Karnivora | Anjing | Hiperkarnivora | AMNH | 12.886 orang | Oligosen | Fotogrametri | Kunjungan koleksi |
| Eoarktos | Karnivora | Ursida | Hipokarnivora | Angkatan Laut AS | Nomor HP 637259 | Oligosen | Pemindaian laser | Sumber Morfo |
| Epiyon haydeni | Karnivora | Anjing | Pemecah tulang | AMNH | 61.462 orang | Miosen | Fotogrametri | Kunjungan koleksi |
| Epikyon saevus | Karnivora | Anjing | Pemecah tulang | Bahasa Indonesia: UF | Wakil Presiden 37265 | Miosen | Permukaan dipindai dan difoto | Kunjungan koleksi |
| Genta genta | Karnivora | Burung Viverridae | Mesokarnivora | MNHN | ZM MO-1997-450 | Permukaan dipindai | Sumber Morfo | |
| Gulo-gulo | Karnivora | Mustelidae | Pemecah tulang | AMNH | Nomor M-182936 | Holosen | Dipindai CT | Sumber Morfo |
| Hemision | Karnivora | Ursida | Hiperkarnivora | Bahasa Indonesia: UF | Wakil Presiden 19908 | Miosen | Permukaan dipindai dan difoto | Kunjungan koleksi |
| Herpes Jawa | Karnivora | Herpestidae | Mesokarnivora | Angkatan Laut AS | 293.772 orang | Holosen | Dipindai CT | Sumber Morfo |
| Burung Hesperocion | Karnivora | Anjing | Mesokarnivora | Angkatan Laut AS | PAL336368 | Oligosen | Fotogrametri | Kunjungan koleksi dan database |
| Anjing hutan anjing hutan | Karnivora | Suku Hyaenidae | Pemecah tulang | Angkatan Laut AS | 182.034 orang | Holosen | Dipindai CT | Sumber Morfo |
| Iktiteria | Karnivora | Suku Hyaenidae | Mesokarnivora | AMNH | KELUARGA 144906 | Miosen | Fotogrametri | Kunjungan koleksi |
| Meles Meles | Karnivora | Mustelidae | Hipokarnivora | AMNH | Nomor telepon M-70603 | Holosen | Dipindai CT | Sumber Morfo |
| Mesocyon sp. | Karnivora | Anjing | Mesokarnivora | Angkatan Laut AS | V 7916 | Oligosen | Fotogrametri | Kunjungan koleksi dan database |
| Mesocyon temnodon | Karnivora | Anjing | Mesokarnivora | Angkatan Laut AS | Nomor 63367 | Oligosen | Fotogrametri | Kunjungan koleksi |
| Metatomarctus | Karnivora | Anjing | Mesokarnivora | Bahasa Indonesia: UF | Wakil Presiden 256998 | Miosen | Permukaan dipindai dan difoto | Kunjungan koleksi dan database |
| Miosin | Karnivora | Burung pemakan serangga | Mesokarnivora | SD NHM | 40.814 | Zaman Eosen | Permukaan dipindai | Dibagikan |
| Mustela frenata | Karnivora | Mustelidae | Hiperkarnivora | Angkatan Laut AS | 95.054 orang | Holosen | Permukaan dipindai | Sumber Morfo |
| Bunga Nandinia binotata | Karnivora | Burung Viverridae | Hipokarnivora | Angkatan Laut AS | 450.440 | Holosen | Dipindai CT | Sumber Morfo |
| Visi Neogale | Karnivora | Mustelidae | Hiperkarnivora | IMNH | Rp 103 | Holosen | Pemindaian laser | Sumber Morfo |
| Osteoborus cyonoides | Karnivora | Anjing | Pemecah tulang | AMNH | Nomor Telepon 61640 | Miosen | Fotogrametri | Kunjungan koleksi |
| Palhyaena | Karnivora | Suku Hyaenidae | Pemecah tulang | AMNH | KELUARGA 144897 | Miosen | Fotogrametri | Kunjungan koleksi |
| Harimau Sumatera | Karnivora | Kucing | Hiperkarnivora | WRAZL | 8.610.108 tahun | Holosen | Fotogrametri | Kunjungan koleksi |
| Paradoxurus hermafroditus | Karnivora | Burung Viverridae | Hipokarnivora | MNHN | ZM AC-A3448 | Holosen | Permukaan dipindai | Sumber Morfo |
| Parahyaena brunea | Karnivora | Suku Hyaenidae | Pemecah tulang | MVZ | 117.842 orang | Holosen | Dipindai CT | Sumber Morfo |
| burung pipit | Karnivora | Anjing | Hipokarnivora | AMNH | Radio FM 8768 | Oligosen | Fotogrametri | Kunjungan koleksi |
| Pliokrokuta | Karnivora | Suku Hyaenidae | Pemecah tulang | Bahasa Indonesia: MNHL | 20.161.768 orang | Zaman Pliosen | Permukaan dipindai | Dibagikan |
| Pliosen | Karnivora | Amphicyonidae | Pemecah tulang | Bahasa Indonesia: UF | Wakil Presiden 24013 | Miosen | Permukaan dipindai dan difoto | Kunjungan koleksi dan database |
| Proailurus | Karnivora | Kucing | Hiperkarnivora | AMNH | 101.931 orang | Miosen | Fotogrametri | Kunjungan koleksi dan database |
| Prokinodikis | Karnivora | Burung pemakan serangga | Mesokarnivora | SD NHM | 42.810 | Zaman Eosen | Permukaan dipindai | Dibagikan |
| Procyon lotor | Kaniformia | Procyonidae | Hipokarnivora | UMMZ | 98.905 tahun | Holosen | Foto | Basis Data |
| Pseudaelurus intrepidus | Karnivora | Kucing | Hiperkarnivora | AMNH | V 124 | Miosen | Fotogrametri | Kunjungan koleksi |
| Pseudaelurus yang validus | Karnivora | Kucing | Hiperkarnivora | AMNH | 62.128 | Miosen | Fotogrametri | Kunjungan koleksi dan database |
| Sepatu Puma Concolor | Karnivora | Kucing | Hiperkarnivora | WRAZL | Holosen | Fotogrametri | Kunjungan koleksi | |
| Stenitis | Karnivora | Mustelidae | Hiperkarnivora | AMNH | FM 25235 | Miosen | Fotogrametri | Kunjungan koleksi dan database |
| Sunkahetanka | Karnivora | Anjing | Hiperkarnivora | YPM | VPPU 013602 | Oligosen | Permukaan dipindai dan difoto | Kunjungan bersama dan koleksi |
| tapokyon | Karnivora | Burung pemakan serangga | Mesokarnivora | SD NHM | 36.000 | Zaman Eosen | Permukaan dipindai | Dibagikan |
| Urosion | Karnivora | Anjing | Mesokarnivora | MUZI | MT1117 | Holosen | Permukaan dipindai | Sumber Morfo |
| Ursus Amerika | Karnivora | Ursida | Hipokarnivora | Universitas Negeri Carolina Utara | 371.168 orang | Holosen | Cahaya terstruktur | Sumber Morfo |
| Vulpavus palustris | Karnivora | Burung pemakan serangga | Mesokarnivora | AMNH | 19.000 | Zaman Eosen | Fotogrametri | Kunjungan koleksi |
| Vulpavus profektus | Karnivora | Burung pemakan serangga | Mesokarnivora | Angkatan Laut AS | PAL362805 | Zaman Eosen | Fotogrametri | Kunjungan koleksi dan database |
| Vulpes vulpes | Karnivora | Anjing | Mesokarnivora | Universitas California, California | Mamalia 15.180 | Holosen | Dipindai CT | Sumber Morfo |
Kami memproduksi model mesh digital 3D dari pindaian asli kami sendiri menggunakan fotogrametri, pemindaian permukaan, atau pemindaian CT dan mengunduh mesh tambahan dari repositori Morphosource (Boyer et al., 2016 ). Tabel 1 melaporkan taksa, sumber mesh, dan data spesimen untuk spesimen dalam penelitian kami. Fotogrametri adalah metode di mana informasi 3D diekstraksi dari foto untuk membuat model 3D (Mikhail et al., 2001 ). Foto diambil melalui kamera Nikon D5600 DSLR dengan lensa 18–55 mm atau lensa AF-S DX Micro NIKKOR 85 mm f 3 5G ED VR (tergantung pada ukuran spesimen) pada meja putar manual dalam kotak lampu LED dengan pencahayaan yang menyebar. Agisoft Metashape 1.8.5 (Agisoft LLC) adalah perangkat lunak yang digunakan untuk membuat model digital dari foto yang diambil oleh kamera dalam prosedur fotogrametri. Pemindaian permukaan dilakukan melalui Pemindai 3D NextEngine atau Pemindai Arctec Spyder, tergantung pada apa yang tersedia di lembaga tempat sampel disurvei. Pemindaian CT dilakukan pada spesimen yang biasanya diunduh dari repositori data tiga dimensi daring Morphosource dan disegmentasi menjadi jaring 3D melalui perangkat lunak 3D Slicer (Pieper et al., 2004 ). Prosedur yang digunakan untuk memperoleh jaring 3D tidak memiliki konsekuensi apa pun untuk penelitian, hanya pada resolusi dan kualitas model permukaan keluaran untuk spesimen tempat kami mengidentifikasi fitur homolog untuk menempatkan landmark.
2.3 Morfometrik Geometri
Morfometrik geometri adalah analisis bentuk menggunakan koordinat landmark dan semilandmark Cartesian (Adams et al., 2004 ; Bookstein, 1991 ; Dryde & Mardia, 1998 ; Perez et al., 2006 ). Koordinat 3D dikumpulkan dari model digital menggunakan alat markup dalam perangkat lunak 3D Slicer (Pieper et al., 2004 ). Landmark dikumpulkan dari mandibula kiri atau mandibula kanan yang dicerminkan ketika yang pertama tidak tersedia atau tidak lengkap. Dua puluh dua landmark tetap diambil dari mandibula seperti yang diuraikan dalam Gambar 2. Landmark ini diturunkan berdasarkan Meloro et al. ( 2015 ), Meloro dan O’Higgins ( 2011 ), dan Prevosti et al. ( 2012 ) untuk menyoroti area utama yang terkait dengan diet dan fungsi.
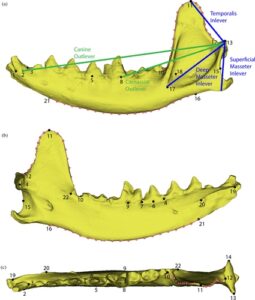
Kurva semilandmark juga digunakan untuk membuat titik-titik berjarak sama yang menelusuri bentuk tepi mandibula di tiga daerah yang penting secara fungsional: satu kurva untuk menangkap kurva rostral prosesus koronoid antara landmark 10 dan 11 dengan 10 titik berjarak sama, satu kurva untuk menangkap kurva kaudal prosesus koronoid antara landmark 11 dan 12 dengan 10 titik berjarak sama, dan satu kurva untuk menangkap sisi ventral ramus horizontal mandibula antara landmark 16 dan 21 dengan 20 titik berjarak sama. Secara total, model 3D memiliki 62 landmark (22 landmark tetap dan 40 semilandmark) (Gambar 2 ).
Analisis morfometri geometrik dilakukan di R (Ihaka & Gentleman, 1996 ) menggunakan rStudio dengan paket R geomorph v4.0.5 (Adams & Otárola-Castillo, 2013 ). Landmark ditumpangkan dengan Procrustes dan diserahkan ke analisis komponen utama (PCA) bentuk untuk memvisualisasikan kelompok landmark mana yang mendorong variasi bentuk. Analisis varians Procrustes berbasis permutasi (ANOVA; Klingenberg & McIntyre, 1998 ) digunakan untuk menguji hubungan bentuk dengan metrik fungsional-morfologi dan biomekanik dari keuntungan mekanis (MA) gigitan temporalis dan masseterik (diukur secara terpisah pada gigi kaninus dan carnassial), rata-rata aritmatika tertimbang mesh (MWAM) tegangan dan regangan Von Mises saat menggigit gigi kaninus dan carnassial secara terpisah (diperkirakan dengan analisis elemen hingga, seperti dijelaskan di bawah), dan analisis kovariansi multivariat faktor tunggal (MANCOVA; Adams & Collyer, 2018 ; Huberty & Petoskey, 2000 ) dari kategori ekologi dan pengelompokan tingkat keluarga taksonomi. Untuk menilai efek filogeni, kuadrat terkecil umum filogenetik (PGLS) juga digunakan untuk menguji hubungan bentuk dengan metrik fungsional morfologi dan biomekanik. Hal ini dipasangkan dengan statistik K umum yang mewakili tingkat sinyal filogenetik. Pohon filogenetik yang dikalibrasi waktu dibuat dari Baskin ( 1998 ), Wozencraft ( 1989 ), Flynn et al. ( 2005 ), dan Spaulding dan Flynn ( 2012 ). Penanggalan didasarkan pada tanggal kemunculan pertama dan terakhir dari Paleobiology Database (Peters & McClenn, 2016 ).
Keuntungan mekanis diambil sebagai proksi untuk gaya gigitan. Keuntungan mekanis adalah rasio panjang tuas dalam (lengan tuas yang menghasilkan gaya, yaitu otot) terhadap panjang tuas luar (lengan tuas tempat gaya tuas dalam disalurkan, yaitu titik gigitan). Regresi metrik seperti keuntungan mekanis ke bentuk Procrustes adalah untuk mengekstrak bagian mandibula yang berhubungan langsung dengan lengan tuas.
2.4 Analisis elemen hingga
Analisis elemen hingga (FEA) diterapkan pada model digital untuk menghitung tegangan dan regangan yang diberikan oleh gigitan simulasi di dua lokasi sepanjang deretan gigi. Analisis elemen hingga mensimulasikan bagaimana suatu objek berperilaku ketika gaya diberikan dengan memperlakukan model sebagai banyak elemen yang terhubung dan terpisah (Korioth et al., 1992 ; Rayfield, 2007 ; Richmond et al., 2005 ; Ross, 2005 ). FEA memecahkan matriks kekakuan di mana gaya adalah masukan dan perpindahan (atau deformasi di setiap simpul), dan kemudian, melalui proses integrasi dalam elemen, tegangan dan regangan diperoleh.
Sebelum melakukan FEA, model diedit dalam Geomagic (3D Systems Corp; Hai, 2007 ) untuk menghilangkan mahkota gigi karena jumlah gigi yang berbeda dan, terutama dalam kasus spesimen fosil, gigi yang hilang (Gill et al., 2014 ; Morales-García et al., 2019 ). Mandibula dan akar gigi yang tersisa diintegrasikan sebagai satu badan karena keduanya kaku dan integrasi ini telah digunakan oleh penulis lain (Gill et al., 2014 ; Morales-García et al., 2019 ; Tseng, 2013 ). Model digital spesimen fosil juga disiapkan untuk FEA mengikuti rekomendasi yang dijelaskan dalam Lautenschlager ( 2016 ) dengan memperbaiki retakan dan mengisi lubang menggunakan Geomagic dan Blender (Brito, 2007 ). Setelah model disiapkan, uji konvergensi pada resolusi elemen menentukan jumlah minimum elemen yang diperlukan untuk menghasilkan hasil tegangan dan regangan yang sama seperti model resolusi yang lebih tinggi (Bright & Gröning, 2011 ; Jones & Wilcox, 2008 ; Maas et al., 2012 ). Jumlah minimum node, permukaan, dan elemen ditemukan rata-rata sebesar 143.000, 223.000, dan 514.000 masing-masing berdasarkan penggunaan elemen tetrahedral untuk jaring.
Elemen model diberi properti elastis isotropik FEA dengan modulus Young sebesar 20 GPa dan rasio Poisson sebesar 0,3 berdasarkan properti rata-rata tulang kortikal mamalia (Currey, 1984 ; Dumont et al., 2005 ) dan studi FEA sebelumnya pada tengkorak Karnivora (Tseng, 2009 ; Tseng et al., 2011 ; Tseng & Flynn, 2015 ). Seluruh mandibula diberi properti ini, mengabaikan akar potensial gigi, karena FEA akan menentukan bagaimana bentuk mandibula yang berbeda akan menangani beban gaya yang berbeda dan mendistribusikan stres (Gill et al., 2014 ; Morales-García et al., 2019 ; Porro et al., 2011 ).
Batasan FEA ditempatkan pada kondilus mandibula dan area paling dorsal dan ventral dari perlekatan otot untuk membuat eksperimen counter-lever (Tseng et al., 2023 ). Batasan multititik ditempatkan pada area ini dengan empat derajat kebebasan yang ditentukan (untuk area perlekatan otot dorsal dan ventral: U1 = U2 = U3 = UR1 = UR2 = UR3 = 0; untuk kondilus mandibula: U1 = U2 = U3 = UR3 = 0; U1 adalah sumbu mediodistal, U2 adalah sumbu dorsovental, dan U3 adalah sumbu sepanjang rahang; U menggambarkan gerakan translasi dan UR menggambarkan gerakan rotasi) (Gill et al., 2014 ). Batasan kondilus mandibula hanya memungkinkan gerakan sendi yang diharapkan seperti yang terjadi in vivo (Gambar 3 ; persegi panjang oranye).
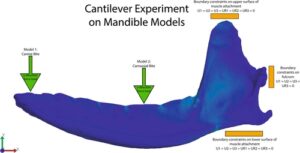
Kami menggunakan dua model, satu di mana beban gaya ekstrinsik ditempatkan pada gigi taring dan yang lain di mana beban gaya ekstrinsik ditempatkan pada gigi carnassial (Gambar 3 ; segitiga hijau). Lokasi-lokasi ini memiliki daya ungkit yang berbeda (rostral versus kaudal) dan fungsi yang berbeda (memperoleh/menaklukkan mangsa dengan gigi taring versus mengunyah/memproses makanan dengan gigi carnassial/molar). Vulpavus profectus , karnivora basal kecil dan mesokarnivora yang disimpulkan, diberi beban 100 N pada setiap area gigi tergantung pada modelnya (gigi taring atau carnassial). Beban 100 N pada Vulpavus ini kemudian diskalakan menurut rasio volume terhadap pangkat 2/3 mandibula lainnya untuk menghasilkan beban yang sebanding untuk model lain dan memungkinkan perbandingan tegangan dan regangan pada mandibula berdasarkan perbedaan bentuk mandibula (Dumont et al., 2005 ; Fortuny et al., 2015 ; Marcé-Nogué et al., 2013 ; Rayfield, 2007 ). MWAM dari tegangan Von Mises dan regangan efektif dicatat dari setiap titik gigitan setiap model (Marcé-Nogué et al., 2017 ; Serrano-Fochs et al., 2015 ).
3 HASIL
3.1 Analisis morfometri geometri
Perbedaan bentuk mandibula yang sebelumnya telah disimpulkan berhubungan dengan fungsi biomekanik mencakup kelengkungan ventral mandibula, orientasi dan ketebalan prosesus koronoid, ketebalan ramus, dan lebar kondilus (Greaves, 1985 ; Van Valkenburgh, 1991 , 1999 , 2007 ; Van Valkenburgh et al., 2004 ). Untuk menilai pola diet dan filogenetik dalam perbedaan bentuk mandibula, tiga sumbu pertama (dari total 53 sumbu) dari morfospace PCA, yang mencakup sekitar 54% dari total varians bentuk, dibangun dengan setiap kelompok ekologi atau filogenetik dibatasi dengan lambung cembung (Gambar 4 ).
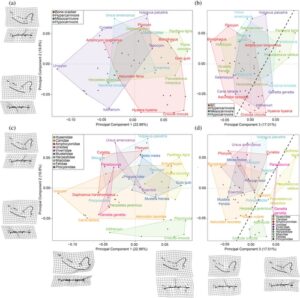
PC1 mewakili 24,96% dari total varian bentuk. Skor positif PC1 mewakili kondilus yang relatif tinggi tetapi sempit, bilah karnasia pendek relatif terhadap area penghancuran molar yang lebih panjang, rostrum yang relatif panjang, fosa maseterik yang relatif kecil, prosesus koronoideus yang relatif pendek, badan ventral mandibula yang lebih melengkung, dan simfisis mandibula horizontal yang relatif panjang. Takson paling ekstrem pada skor ini adalah Urocyon (rubah abu-abu). Skor negatif PC1 mewakili kondilus yang relatif lebar dan terletak di rostral, bilah karnasia panjang relatif terhadap area penghancuran molar yang lebih kecil, rostrum yang relatif pendek, fosa maseterik yang relatif rostral, prosesus koronoideus yang relatif besar dan terletak distal, badan ventral mandibula yang relatif lurus, dan simfisis mandibula vertikal yang relatif besar dengan gigi taring yang besar. Takson paling ekstrem pada skor ini adalah Puma (singa gunung).
PC2 mewakili 17,32% dari total varians bentuk. Skor negatif PC2 adalah bilah carnassial yang panjang, gigi geraham depan keempat yang lebih kaudal, simfisis mandibula yang lebih rostral, dan fosa maseterik yang lebih ventral; takson yang paling ekstrem adalah Crocuta (hyena tutul). Skor positif PC2 mewakili gigi geraham depan keempat dan bilah carnassial yang lebih rostral, simfisis mandibula yang lebih kaudal; takson yang paling ekstrem adalah Vulpavus .
PC3 mewakili 12,19% dari total varian bentuk. Skor negatif PC3 mewakili ramus horizontal yang lebih bersudut, prosesus koronoideus yang lebih tinggi dan lebih curam, premolar keempat dan carnassial yang lebih rostral, masseter yang lebih rostral, dan prosesus koronoideus yang curam; takson yang paling ekstrem adalah Borophagus . Skor positif PC3 adalah ramus horizontal yang lebih lurus, area penghancuran molar yang lebih kecil, prosesus koronoideus yang lebih rendah dan kurang curam, dan masseter yang lebih rostral; takson yang paling ekstrem adalah Panthera (khususnya harimau).
PC pertama tampaknya merupakan kontras pola makan antara mesokarnivora dan morfologi hiperkarnivora dan pemecah tulang yang terspesialisasi, dengan beberapa tumpang tindih di tengah (Gambar 4 ). Ada sedikit pemisahan antara kategori pola makan yang tampak pada PC2 dan PC3. Mesokarnivora menempati bagian morfospace yang lebih besar daripada kategori pola makan lainnya, dengan pemecah tulang memiliki penyebaran terbesar kedua, diikuti oleh hiperkarnivora lainnya. Tumpang tindih pemecah tulang dan mesokarnivora sebagian besar didorong oleh amfikionida (kelompok karnivora mirip canid yang telah punah yang secara sehari-hari disebut sebagai “anjing beruang”) di mana beberapa spesies telah disimpulkan sebagai pemecah tulang (misalnya, Amphicyon ). Hipokarnivora muncul di tengah antara tiga kelompok lainnya. Hipokarnivora umumnya bersarang di tengah semua kelompok lainnya, di mana mereka saling tumpang tindih, sebagian besar di sisi negatif PC1, ujung positif PC2, dan berpusat pada PC3. Ada tumpang tindih yang lebih tinggi antara mesokarnivora dan hipokarnivora ketika melihat PC2 dan PC3 (Gambar 4b ).
Pengelompokan filogenetik paling jelas terlihat pada PC2 dan PC3 (Gambar 4d ) di mana Hyaenidae, Felidae, dan Viverridae ditampilkan. Canidae menempati area terbesar dalam plot, dan Caninae (yaitu, Canis , Vulpes , Urocyon , dan Aenocyon ) berbeda dari canid lainnya (yaitu, Hersperocyoninae seperti Hesperocyon dan Borophaginae seperti Epicyon ). Kombinasi PC2 dan PC3 menunjukkan pemisahan antara Feliformia dan Caniformia (Gambar 4b,d ). Untuk PC pertama, dua spesies dari genus Vulpavus , genus karnivora yang lebih basal, biasanya berbeda dari karnivora lainnya, sedangkan Tapocyon sangat mirip dengan mustelida dan canid, Miocyon dengan amphicyonida, dan Procynodictis dengan canid dan viverrid. Perlu dicatat bahwa Herpestes berada pada garis yang memisahkan Feliformia dan Caniformia. Pengambilan sampel Herpestidae yang lebih besar atau feliform lain yang lebih kecil atau secara morfologi lebih umum, seperti Viverridae dan Eupleridae, mungkin membuat pemisahan ini kurang jelas pada PC ini.
3.2 Morfologi fungsional: keuntungan mekanis
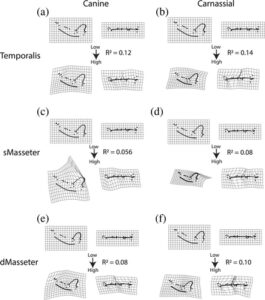
Perkiraan keuntungan mekanis diregresikan ke jarak Procrustes dari masing-masing taksa untuk menentukan daya penjelasan yang dimiliki setiap metrik fungsi pada bentuk mandibula (Tabel 2 ). Nilai R 2 tidak lebih besar dari 0,14 untuk metrik apa pun, dengan rata-rata 0,10, dan semuanya terbukti signifikan ( nilai- p <0,05). Nilai tertinggi adalah keuntungan mekanis temporalis dan kaninus ( R 2 = 0,20, nilai- p = 0,001) dan nilai terendah adalah keuntungan mekanis masseter superfisial dan kaninus ( R 2 = 0,06, nilai- p = 0,004). Daya penjelasan untuk sebagian besar metrik ditingkatkan di bawah MANCOVA ketika filogeni berkovariasi dengan bentuk.
| Kategori atau metrik fungsional | p – Nilai kategori atau metrik pada bentuk Procrustes | R 2 kategori atau metrik ke bentuk Procrustes | Tiga komponen utama berkorelasi tertinggi dari bentuk ke metrik | p – Nilai kategori atau metrik pada bentuk Procrustes (PGLS) | R 2 kategori atau metrik pada bentuk Procrustes (PGLS) |
|---|---|---|---|---|---|
| Ekologi | 0,001 | 0.18 | Bahasa Indonesia | 0,168 | 0,09 |
| Keluarga | 0,001 | 0.38 | Bahasa Indonesia | 0.707 | 0.18 |
| Subordo | 0,001 | 0.14 | Bahasa Indonesia | 0.233 | 0,09 |
| MA Anjing—Temporalis | 0,001 | 0.2 | 1,4,7 | 0,031 | 0,06 |
| MA Carnassial—Temporalis | 0,001 | 0.14 | 1,2,4 | 0,044 tahun | 0,05 |
| MA Canine—Masseter Superfisial | 0,004 tahun | 0,056 tahun | 1,7,4 | 0.109 | 0,05 |
| MA Carnassial — Masseter Dangkal | 0,001 | 0,08 | 1,7,4 | 0,158 | 0,05 |
| MA Canine—Masseter Dalam | 0,001 | 0,08 | 1,3,2 | 0,050 | 0,02 |
| MA Carnassial—Masseter Dalam | 0,001 | 0.10 | 1,3,2 | 0,081 tahun | 0,02 |
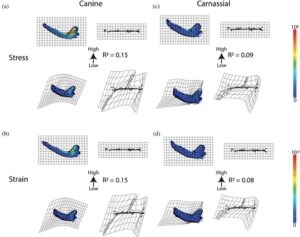
| FEA—stres pada Anjing | 0,001 | 0,15 | 1,5,4 | 0,035 | 0,05 |
| FEA—strain pada Anjing | 0,001 | 0,15 | 1,5,4 | 0,035 | 0,05 |
| FEA—stres di Carnassial | 0,001 | 0,09 | 1,2,6 | 0,068 tahun | 0,07 |
| FEA—strain di Carnassial | 0,001 | 0,08 | 1,2,6 | 0,068 tahun | 0,06 |
Singkatan: MA, keuntungan mekanis.
Untuk PGLS, semua metrik menunjukkan pengaruh filogeni dalam korelasinya dengan bentuk karena semua nilai R 2 menurun dan beberapa menjadi tidak signifikan ( nilai- p >0,05). Satu-satunya metrik yang masih mempertahankan nilai signifikan adalah keuntungan mekanis temporalis dan kaninus ( R 2 = 0,06, nilai- p = 0,031), temporalis dan carnassial ( R 2 = 0,05, nilai- p = 0,044), dan masseter dalam dan kaninus ( R 2 = 0,02, nilai- p = 0,05). Statistik- K umum pada derajat sinyal filogenetik pada variabel bentuk adalah 0,3276, yang menunjukkan beberapa efek filogeni pada bentuk mandibula.
Ketika skor komponen utama diregresikan ke metrik fungsional, hampir semua metrik paling berkorelasi dengan PC1 (Tabel 2 ). PC2 sangat berkorelasi dengan keuntungan mekanis temporalis di carnassial dan kedua pengukuran masseter dalam, dan stres carnassial. PC3 hanya sangat berkorelasi dengan metrik keuntungan mekanis masseter dalam (Tabel 2 ). PC yang lebih rendah yang juga lebih berkorelasi dengan metrik termasuk PC4 (pengukuran keuntungan mekanis sebagian besar kecuali dengan masseter dalam), PC5 (stres dan regangan kaninus), PC6 (stres dan regangan carnassial), dan PC7 (keuntungan mekanis temporalis dan kaninus dan kedua pengukuran masseter superfisial). PC4, PC5, PC6, dan PC7 mewakili 9,22%, 6,2%, 4,27%, dan 3,53% dari total varians bentuk dalam urutan tersebut dan berjumlah 23,22% dari total varians bentuk (dengan PC 1–3 menghasilkan sekitar 77% dari total varians bentuk).
Deformasi dalam bentuk yang terkait dengan masing-masing parameter fungsional ini divisualisasikan dengan splines pelat tipis, di sini ditunjukkan sebagai bentuk yang terkait dengan nilai tinggi setiap metrik (Gambar 5 ). Parameter fungsional dengan daya penjelasan tertinggi, keuntungan mekanis temporalis dan carnassial melibatkan pemendekan mandibula, memiliki orientasi simfisis mandibula yang lebih vertikal, pengurangan area penghancuran mandibula, dan pelebaran kondilus (Gambar 5b dan S2 ). Karakteristik ini biasanya terkait dengan derajat hiperkarnivora yang lebih besar (misalnya, pengurangan area penghancuran mandibula) dan jumlah tekanan yang lebih tinggi pada mandibula (misalnya, kondilus yang lebih lebar, simfisis mandibula yang lebih vertikal), tetapi nilai keuntungan mekanis terendah berasal dari Epicyon saevus dan nilai keuntungan mekanis tertinggi berasal dari Eoarctos .
3.3 Morfologi fungsional: analisis elemen hingga
Analisis varians berbasis permutasi (ANOVA) menunjukkan bahwa stres dan regangan pada mandibula berbeda secara signifikan di antara mesokarnivora, hipokarnivora, hiperkarnivora, dan pemecah tulang dalam karnivora (tiga derajat kebebasan dan nilai- p >0,01 untuk stres dan regangan beban anjing dan =0,02 untuk stres dan regangan beban karnasial). Rata-rata, mesokarnivora memiliki stres dan regangan tertinggi dari beban anjing dan beban karnasial (Tabel 3 ). Stres dan regangan rata-rata di antara hipokarnivora, hiperkarnivora, dan pemecah tulang lebih dekat satu sama lain, tetapi hiperkarnivora dan pemecah tulang memiliki stres dan regangan rata-rata yang lebih rendah dari beban anjing dan beban karnasial. Dalam hal perbedaan stres dan regangan antara klade yang berbeda, “Miacidae” dan Viverridae memiliki stres dan regangan tertinggi dari beban anjing, diikuti oleh Amphicyonidae, Canidae, Procyonidae, Herpestidae, Ursidae, Mustelidae, Hyaenidae, dan Felidae. “Miacidae” dan Amphicyonidae memiliki stres dan regangan tertinggi dari beban carnassial, diikuti oleh Viverridae, Procyonidae, Ursidae, Canidae, Mustelidae, Felidae, Herpestidae, dan Hyaenidae. Di antara dua subordo, caniform memiliki stres dan regangan rata-rata yang lebih tinggi daripada feliform.
| Ekologi | Stres anjing (Pa) | Strain anjing | Tekanan karnasial (Pa) | Strain Carnassial |
|---|---|---|---|---|
| Mesokarnivora | 1.58E+07 | 1.03E−03 | 4.02E+06 | 2.65E−04 |
| Hipokarnivora | 1.16E+07 | Nomor 7.56E−04 | 2.81E+06 | 1.83E−04 |
| Hiperkarnivora | 1.08E+07 | 7.01E−04 | 2.48E+06 | 1.61E−04 |
| Pemecah tulang | 1.07E+07 | 6.98E−04 | 2.66E+06 | 1.73E−04 |
| Keluarga | ||||
| Burung Viverridae | 1.56E+07 | 1.01E−03 | Nomor telepon 3.24E+06 | 2.11E−04 |
| Burung pemakan serangga | 1.55E+07 | 1.00E−03 | Tanggal 5.29E+06 | 3.56E−04 |
| Amphicyonidae | 1.40E+07 | 9.09E−04 | 3.84E+06 | 2.50E−04 |
| Anjing | 1.33E+07 | 8.65E−04 | Nomor telepon 3.21E+06 | 2.09E−04 |
| Procyonidae | 1.33E+07 | 8.62E−04 | Tanggal 3.29E+06 | 2.14E−04 |
| Herpestidae | Tanggal 1.19E+07 | Nomor 7.72E−04 | 2.03E+06 | 1.32E−04 |
| Ursida | 1.14E+07 | Nomor 7.44E−04 | Nomor telepon 3.34E+06 | 2.17E−04 |
| Mustelidae | 1.07E+07 | Nomor 6.93E−04 | 2.36E+06 | 1.54E−04 |
| Suku Hyaenidae | 1.03E+07 | 6.68E−04 | 1.87E+06 | Nomor 1.22E−04 |
| Kucing | 9.47E+06 | 6.15E−04 | 2.04E+06 | 1.33E−04 |
Catatan : Daftar rata-rata kelompok stres dan regangan MWAM Von Mises pada mandibula akibat beban anjing dan karnivora dengan kelompok berdasarkan ekologi atau klade. Stres dan regangan diperoleh dari beban 100 N yang diskalakan. Perlu dicatat bahwa Procyonidae dan Herpestidae keduanya diwakili oleh 1 takson.
Bahasa Indonesia : Ketika diregresikan ke data bentuk yang selaras dengan Procrustes, metrik tegangan dan regangan secara signifikan hampir sama dalam kemampuannya untuk menjelaskan variasi bentuk mandibula (Tabel 2 ). Nilai tegangan dan regangan R2 signifikan dan sebanding dengan nilai R2 dari keuntungan mekanis temporalis (Tabel 2 ). Tegangan dan regangan dari beban anjing lebih baik dalam menjelaskan bentuk daripada beban karnasial. MANCOVA dari interaksi antara filogeni atau ekologi meningkatkan daya penjelasan. Mirip dengan pengukuran keuntungan mekanis, PGLS menunjukkan penurunan nilai R2 dari pengukuran tegangan dan regangan yang menunjukkan pengaruh filogeni. Tegangan dan regangan anjing memiliki signifikan ( nilai- p = 0,035) R2 = 0,05, tetapi tegangan dan regangan karnasial memiliki tidak signifikan ( nilai-p = 0,07 dan 0,06 , berturut-turut. Namun, pengukuran tegangan dan regangan anjing masih sebanding dengan pengukuran keuntungan mekanis.
Ketika tekanan dan regangan pada mandibula berkurang, mandibula menjadi lurus dan memendek, area penghancuran molar berkurang, kondilus berada pada posisi yang lebih ventral dan menjadi lebih lebar, lingkar gigi kaninus meningkat, area simfisis mandibula meningkat dan menjadi lebih berorientasi vertikal, dan area perlekatan otot bergerak lebih ke arah rostral (Gambar 6 ). Adcrocuta , pemecah tulang dari Hyaenidae, dan Pseudaelurus validus , hiperkarnivora dari Felidae, memiliki tekanan dan regangan terendah dari beban gigi kaninus dan carnassial. Vulpes dan Miocyon , mesokarnivora dari Canidae dan “Miacidae”, memiliki tekanan dan regangan tertinggi dari beban gigi kaninus, dan Amphicyon dan Miocyon , yang sebelumnya merupakan Amphicyonidae, memiliki tekanan dan regangan tertinggi dari beban carnassial. Nilai tegangan dan regangan Miocyon sangat tinggi dan mungkin disebabkan oleh kesalahan perbaikan yang dilakukan pada model 3D asli spesimen tersebut.
4 DISKUSI
4.1 Pola tekanan dan regangan pada mandibula
Mesokarnivora cenderung, secara rata-rata, memiliki nilai stres dan regangan yang lebih tinggi dari beban mandibula daripada ekologi karnivora khusus seperti hiperkarnivora dan hiperkarnivora pemecah tulang. Tingkat karnivora tampaknya berkorelasi dengan stres dan regangan yang lebih rendah kecuali dengan hipokarnivora dan beban karnasial (Tabel 3 ) yang cenderung memiliki stres dan regangan yang lebih rendah daripada hiperkarnivora tetapi tidak pada pemecah tulang. Mesokarnivora beradaptasi dengan pola makan generalis dengan mangsa yang jauh lebih kecil daripada mereka, dengan mandibula yang lebih panjang, seringkali lebih tipis di arah dorsoventral, untuk menangkap mangsa yang tidak akan melibatkan banyak tekanan komparatif pada mandibula dari mangsa yang berjuang dibandingkan dengan hiperkarnivora (Christiansen, 2008 ; Christiansen & Adolfssen, 2005 ; Christiansen & Wroe, 2007 ; Davis, 2014 ; Martin, 1980 ; Meloro & O’Higgins, 2011 ; Roemer et al., 2009 ; Van Valkenburgh, 2007 ). Dalam sampel studi ini, bentuk mandibula mesokarnivora rata-rata mencakup kelengkungan yang lebih besar pada sisi ventral mandibula, simfisis mandibula yang lebih tipis dan lebih sejajar dengan rostro-kaudal, dan prosesus koronoid yang lebih pendek sehingga menghasilkan lengan pengungkit yang lebih kecil (Gambar 6 ). Ini semua adalah adaptasi yang dimaksudkan untuk meningkatkan jangkauan mandibula untuk menangkap mangsa dan menutup rahang dengan cepat, tetapi dengan mengorbankan kekokohan dan kekuatan mandibula (Christiansen, 2008 ; Meloro et al., 2011 ; Piras et al., 2013 ; Prevosti et al., 2012 ; Van Valkenburgh, 2007 ).
Hiperkarnivora, termasuk hiperkarnivora pemecah tulang, memiliki rahang yang lebih kuat dengan, rata-rata, tekanan dan regangan yang lebih rendah daripada mesokarnivora. Kemampuan menahan kekuatan tinggi dan mengurangi tekanan dan regangan diperlukan bagi hiperkarnivora untuk melawan mangsa yang meronta yang ukurannya sama atau lebih besar dari karnivora (Echarri et al., 2017 ; Meloro et al., 2011 ; Prevosti et al., 2012 ; Tseng et al., 2023 ; Tseng & Flynn, 2018 ). Dalam penelitian ini, bentuk mandibula hiperkarnivora rata-rata mencakup simfisis mandibula yang lebih tebal, lebih sejajar dorsoventral yang digabungkan dengan gigi taring yang lebih besar, sisi ventral mandibula yang lebih lurus (lebih demikian pada hiperkarnivora) dengan hanya melengkung ke arah medial (yang terakhir lebih jelas pada pemecah tulang), kondilus yang lebih besar, dan prosesus koronoid yang lebih tinggi dan fosa maseterik yang lebih rostral (dan untuk pemecah tulang lebih ventral) untuk lengan pengungkit yang lebih panjang (Gambar 7 ). Pengecualian untuk generalisasi ini adalah bahwa pemecah tulang memiliki perpindahan yang nyata antara posisi ramus asenden dan ramus horizontal mandibula (Gambar 7 ). Perpindahan antara dua bagian mandibula meningkatkan lengan pengungkit temporalis dan menggeser kondilus dan deretan gigi. Bentuk mandibula menghasilkan gaya gigitan yang lebih kuat dengan lengan dalam yang lebih panjang dari otot temporalis dan masseter, serta rahang yang lebih kuat dan lebih tahan terhadap gaya gigitan yang lebih kuat dan gaya dari mangsa yang berontak (ramus horizontal yang lebih lurus dan lebih pendek menjaga kekuatan mandibula tetap tinggi di berbagai bagian mandibula).
Bentuk simfisis mandibula dan ukuran gigi taring berhubungan dengan kekuatan mandibula saat melawan mangsa yang berontak saat menangkap mangsa. Pada hiperkarnivora seperti felid dan beberapa mustelida, gigi taring besar dan simfisis mandibula yang besar menambah kekuatan yang berhubungan dengan penangkapan mangsa yang ukurannya sama atau lebih besar (Christiansen, 2008 ; Davis, 2014 ; Van Valkenburgh, 2007 ). Korelasi yang lebih besar antara stres dan regangan gigi taring pada bentuk mandibula versus stres dan regangan karnasial menekankan pentingnya area gigi taring untuk hiperkarnivora dalam bentuk mandibula. Ada kemungkinan bahwa korelasi antara metrik stres dan regangan dan bentuk mandibula lebih berkaitan dengan ukuran mangsa relatif daripada pola makan.
Melihat kelompok keluarga, beberapa pola dapat dikenali antara stres dan regangan dan mesokarnivora versus hiperkarnivora (Tabel 3 ). Kelompok yang umumnya mesokarnivora, seperti Viverridae, memiliki stres dan regangan yang lebih tinggi daripada kelompok yang mencakup hiperkarnivora, seperti Felidae. Ketika stres dipetakan ke pohon filogenetik di bawah model gerak Brown evolusi, kelompok taksonomi yang menunjukkan penurunan stres dan regangan anjing pada mandibula saat derajat karnivora meningkat terutama tampak pada Canidae ( Hesperocyon dibandingkan dengan taksa terminal Hesperocyoninae dan Borophaginae) dan Hyaenidae ( Icitherium vs. taksa terminal) (Gambar 8a ). Pengecualian untuk yang pertama adalah anggota subfamili Caninae (yaitu, Vulpes , Urocyon , Canis latrans , dan Aenocyon ) yang dikelompokkan bersama dan sebagian besar adalah mesokarnivora (meskipun Canis latrans , anjing hutan, dianggap sebagai mesokarnivora, dengan tidak adanya Canis lupus , serigala abu-abu, atau hiperkarnivora lainnya, ia dapat menjadi lebih hiperkarnivora (Avrin et al., 2023 )). Pola stres dan ketegangan yang berulang dan evolusi menjadi karnivora di Canidae jelas karena kekayaan catatan fosil Amerika Utara untuk Canidae (Balisi et al., 2018 ; Van Valkenburgh, 1991 ; Van Valkenburgh et al., 2004 ). Pola penurunan stres dan tekanan lebih sulit dilihat pada kelompok yang cepat berubah menjadi hiperkarnivora, seperti Felidae (anggota basal, Proailurus , sudah menjadi hiperkarnivora dan satu-satunya takson saudara bagi Proailurus dalam penelitian ini adalah Nandinia , mesokarnivora/hipokarnivora dari Nandiniidae) (Christiansen, 2008 ; Hunt Jr, 1998 ) atau kelompok yang cepat memperkuat mandibula mereka (misalnya, Hyaenidae pada Gambar 8b ).
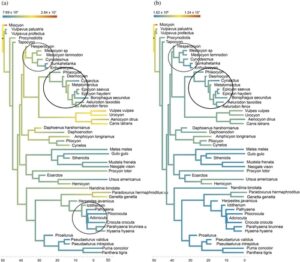
Tren penurunan stres dan regangan dari mesokarnivora ke karnivora khusus umumnya berlaku untuk beban anjing dan carnassial. Pengecualiannya adalah Amphicyonidae, yang memiliki pengukuran stres dan regangan yang lebih tinggi dari beban carnassial (Tabel 3 ). Ini mungkin karena amphicyonida memiliki carnassial yang ditempatkan lebih rostral dan molar postcarnassial yang lebih besar. Setelah menempatkan beban carnassial ke molar kedua yang lebih kaudal, stres dan regangan pada mandibula menjadi lebih sebanding dengan pemecah tulang dan hiperkarnivora lainnya. Pengecualian lain adalah dalam Hyaenidae, karena Ictitherium telah mengalami stres dan regangan rendah dari beban carnassial. Pengecualian dengan Hyaenidae bisa menjadi kasus yang mirip dengan Felidae dari suatu famili yang dengan cepat memperoleh sifat-sifat mandibula yang lebih kuat untuk area carnassial.
4.2 Filogeni versus ekologi dan korelasinya dengan bentuk
Sumbu utama bentuk mandibula (PC1) tampak berkorelasi dengan ekologi dan fungsi dan umumnya memisahkan mesokarnivora dari hiperkarnivora (Gambar 4 ). Ada contoh taksa yang tumpang tindih antara mesokarnivora dan hiperkarnivora/pemecah tulang di PC1, dengan terutama Amphicyon dan Herpestes . Penempatan Amphicyon dalam morfospace dan ketidakmampuan untuk dipisahkan mungkin merupakan bukti kesulitan dalam menentukan ekologi dan pola makannya dalam literatur sebelumnya (Sorkin, 2006 ). Penempatan Herpestes mungkin filogenetik, karena sering kali dekat dengan Viverridae (yang terdiri dari mesokarnivora dan hipokarnivora) yang merupakan kelompok saudara dari kelompok taksonomi Herpestidae yang lebih besar, Herpestoidea, yang dibuatnya dengan Hyaenidae. Pemecah tulang memiliki tumpang tindih yang nyata dengan hiperkarnivora lainnya, tetapi pemecah tulang dengan bentuk yang sama dan filogeni yang lebih dekat cenderung mengelompok di sekitar satu sama lain pada kombinasi PC tanpa PC1. Hipokarnivora juga tumpang tindih di area tertentu dengan mesokarnivora, yang mungkin dikaitkan dengan aspek bentuk mandibula yang terkait dengan area penghancuran molar. Namun, hipokarnivora juga tumpang tindih dengan pemecah tulang dan hiperkarnivora, yang mungkin terkait dengan area perlekatan otot mandibula yang lebih besar dan kedalaman mandibula. Selain itu, selama regresi setiap metrik fungsional ke komponen utama, PC1 secara konsisten paling berkorelasi dengan metrik fungsional, dibandingkan dengan PC lainnya (Tabel 2 ).
Data regresi menunjukkan bahwa ada korelasi kuat antara bentuk dan pengelompokan taksonomi pada tingkat famili ( R 2 = 0,38, nilai- p = 0,001) tetapi korelasi yang lebih lemah pada tingkat subordo ( R 2 = 0,14, nilai- p = 0,001). Plot PCA telah menunjukkan area morfospace yang berbeda yang ditempati oleh famili yang berbeda seperti Felidae dan Hyaenidae (Gambar 4b,d ). Namun, famili kaniform cenderung tumpang tindih satu sama lain pada sumbu morfospace PC pertama. Korelasi antara bentuk dan famili dan apa yang terlihat dari PCA dapat berarti bahwa morfologi mandibula tertentu berevolusi pada tingkat famili. Beberapa morfologi ini dapat dikaitkan dengan contoh pengurangan baris gigi dan bentuk serta penempatan karnasia. Penelitian sebelumnya telah menunjukkan bahwa pada tingkat taksonomi famili, karnivora dengan cepat terdiversifikasi menjadi morfologi gigi yang berbeda yang bertahan di seluruh famili (Hopkins et al., 2022 ). Pengecualian untuk morfologi penguncian ini mungkin adalah karnivora Amerika Utara yang berevolusi selama “celah kucing” di Miosen, seperti Enhydrocyon , yang memiliki tengkorak yang lebih mirip kucing dan mandibula yang plotnya dekat dengan kucing pada sumbu PC pertama (Gambar 4 ). Konvergensi ekologis yang serupa dalam bentuk yang melampaui batasan familial filogenetik juga dapat dilihat pada spesies canid borophagine selanjutnya seperti Aelurodon , Epicyon , dan Borophagus , yang memiliki morfologi yang lebih mirip hyaenid di mandibula mereka.
5 KESIMPULAN
Sementara filogeni sangat berkorelasi dengan variasi bentuk, sumbu utama variasi bentuk pada mandibula karnivora yang direpresentasikan oleh posisi dan lebar kondilus, panjang bilah carnassial, panjang dan kelengkungan mandibula, ukuran area perlekatan otot, dan bentuk simfisis mandibula secara nyata berkorelasi dengan ekologi fungsional berdasarkan regresi metrik fungsional dan pemisahan mesokarnivora dari hiperkarnivora dan pemecah tulang yang lebih terspesialisasi. Selain itu, analisis elemen hingga menunjukkan rata-rata tekanan dan regangan yang lebih tinggi pada mandibula mesokarnivora daripada pada ekologi yang lebih terspesialisasi seperti hiperkarnivora dan pemecah tulang.
Pekerjaan di masa mendatang mengenai hal ini akan mencakup penjelajahan apakah pola yang terlihat dalam kumpulan data ini masih dapat diamati dalam kumpulan data dua dimensi. Untuk yang terakhir, mungkin menarik untuk melihat kelompok taksonomi tertentu yang memiliki tren perubahan ekologi fungsional yang lebih jelas pada tingkat famili (misalnya, Canidae atau subfamili khusus Canidae seperti Hesperocyoninae dan Borophaginae). Hal menarik lainnya adalah melihat bagaimana aspek bentuk, kinerja fungsional, dan ekologi makanan berubah bersamaan satu sama lain (misalnya, apakah mereka berubah pada tingkat yang sama bersamaan satu sama lain atau apakah salah satunya berubah lebih cepat daripada yang lain pada “lag”).